
Насекомое с неполным превращением (с прямым развитием) проходит в своём развитии три стадии: яйцо - личинка - взрослое насекомое (имаго) .
Обрати внимание!
Отряды насекомых с неполным превращением: прямокрылые, равнокрылые, клопы (полужесткокрылые), стрекозы, тараканы, богомолы, подёнки, веснянки, уховёртки, вши.
Личинки очень похожи на родителей и отличаются от взрослых насекомых только меньшей величиной, отсутствием крыльев и неразвитостью половой системы.
Личинки несколько раз линяют, растут с каждой линькой и все больше и больше становятся похожими на взрослых насекомых. Со временем у них полностью формируются крылья и они становятся половозрелыми.
Отряд Стрекозы
Стрекозы - это хорошо известные насекомые с длинным стройным телом и двумя парами сильных прозрачных крыльев.
Стрекозы (особенно крупные) отличаются очень быстрым и маневренным полетом. Они - хищники, ловящие насекомых (мух, комаров, мелких бабочек) на лету. У стрекоз большие сложные глаза, обеспечивающие почти круговой обзор, и длинные ноги, усаженные жесткими волосками.
Личинки стрекоз малоподвижны, живут в прудах, озерах, канавах с водой, медленно текущих реках. Они тоже хищники и хватают проплывающих мимо рачков, личинок других насекомых, головастиков и мальков рыб при помощи способной выбрасываться вперёд нижней губы, которая называется маской.
Отряд Прямокрылые
К этой группе относятся саранча, кузнечики, сверчки и медведки. У них две пары крыльев (передние более плотные, чем задние), у многих задние конечности прыгательные, ротовой аппарат грызущий. Некоторые виды саранчи хорошо летают. Саранча питается растениями, среди кузнечиков есть как растительноядные виды, так и хищники, сверчки - всеядные.
Отряд Равнокрылые
К равнокрылым принадлежат цикады и тли. Ротовой аппарат у них колюще-сосущего типа, а крылья обычно складываются крышей («домиком»). Питаются равнокрылые соком растений.
Цикады - крупные (длиной до 7 см) дневные насекомые и известны тем, что способны издавать очень громкие звуки при помощи особых органов, расположенных снизу у основания брюшка.
Тли - мелкие насекомые длиной 0,5-6 мм. Среди них есть как крылатые, так и бескрылые формы. Многие вредят культурным растениям.
Отряд Клопы, или Полужесткокрылые
Представителей этой группы называют так потому, что их передние крылья (надкрылья) спереди плотные, а сзади мягкие. Вторая пара крыльев лежит под первой. Именно при помощи второй пары крыльев клопы могут летать. У некоторых, например у постельного клопа, крылья отсутствуют. Ротовой аппарат у клопов колюще-сосущий. Среди клопов есть виды, питающиеся соками растений, есть хищники и кровососы (постельный клоп).
Рис. 1. Развитие насекомого с неполным превращениемУ насекомых существует два основных типа развития, в соответствии с которыми их подразделяют на два крупных отдела.
Развитие с неполным превращением проходит в три стадии: яйцо - личинка - имаго (взрослое насекомое), не сопровождаясь резкими изменениями (рис. 1 ). Личинки по строению подобны взрослым насекомым и превращаются в них сразу после окончания развития и последней линьки.
Развитие с полным превращением проходит в четыре стадии: яйцо - личинка - куколка - имаго. Личинки резко отличаются от имаго, поэтому, окончив развитие, они превращаются в покоящуюся стадию куколки, в которой происходит преобразование организации тела от личиночной до имагинальной.
Строение и формы яиц
Яйца насекомых богаты желтком и покрыты двумя оболочками. Они разнообразны по размерам, форме и окраске. Размеры яиц обычно составляют от десятых долей миллиметра до нескольких миллиметров. По форме яйца бывают овальными (многие жуки, бабочки, мухи), удлиненными (кузнечики и саранчовые), шаровидными (клопы-черепашки), полушаровидными (бабочки-совки), бочонковидными (клопы-щитники), бутылковидными (дневные бабочки), стебельчатыми (златоглазки) (рис. 2 ).
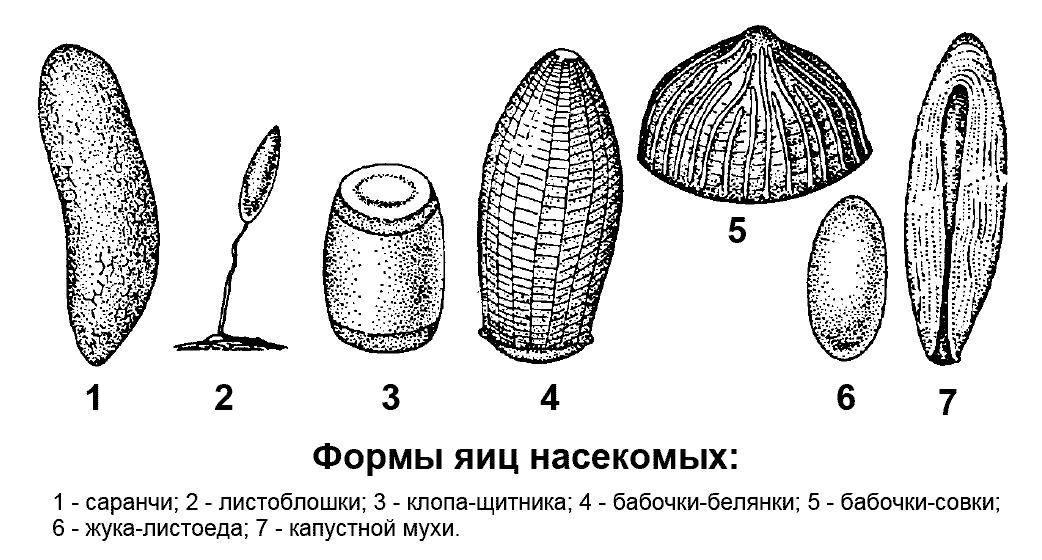 Рис. 2. Формы яиц насекомых
Рис. 2. Формы яиц насекомых Самки различных насекомых откладывают яйца в места, пригодные для дальнейшего развития личинок. Плодовитость насекомых (количество яиц, откладываемое самкой в течение жизни) разнообразна. У некоторых видов она невелика - порядка нескольких десятков яиц, у большинства около нескольких сотен, у ряда видов - несколько тысяч. Плодовитость половозрелых самок общественных насекомых достигает нескольких миллионов яиц. У некоторых насекомых самки откладывают яйца одиночно или врассыпную, однако большинство откладывает их компактными группами - яйцекладками.
Строение и типы личинок
Основные функции личиночной стадии - питание, рост, развитие. В развитии насекомых личинка - единственная растущая стадия. Ее размеры могут увеличиваться в десятки и сотни раз. Поэтому личинки несколько раз проходят линьку, сбрасывая старую кутикулу в виде личиночной шкурки и одновременно образуя новую, рассчитанную на дальнейший рост.
Имагообразные линийки, или нимфы, - личинки насекомых с неполным превращением. Эти личинки сходны по строению со взрослыми насекомыми. Они имеют на грудном отделе три пары хорошо развитых ног, фасеточные глаза, ротовой аппарат, подобный аппарату имаго, хорошо развитые антенны. У них развиваются наружные зачатки крыльев. Обычно имагообразные личинки сходны с имаго также по образу жизни и характеру питания. У вредителей с неполным превращением личинки и имаго всегда повреждают растения сходным образом. Некоторое исключение составляют водноживущие личинки стрекоз и поденок, имеющие органы водного дыхания - трахейные жабры. Их иногда называют наядами.
Личинок насекомых с полным превращением иногда называют истинными . Эти личинки могут иметь различное количество конечностей, развитых слабее, чем у взрослых насекомых; либо конечности отсутствуют. Личинки лишены фасеточных глаз, органами зрения служат простые глазки. У них отсутствуют наружные зачатки крыльев, антенны короткие или неразвиты, ротовой аппарат чаще всего грызущий, он может быть совершенно иного строения, нежели у имаго. Личинки насекомых с полным превращением могут резко отличаться от имаго по образу жизни и характеру питания. У вредителей с полным превращением вред может наносить либо личинка, либо имаго, или насекомые в обеих стадиях развития.
У насекомых с полным превращением выделяют три типа личинок (рис. 3 ).
 Рис. 3. Типы личинок насекомых с полным превращением
Рис. 3. Типы личинок насекомых с полным превращением Камподеовидные личинки (от лат. названия первичнобескрылого насекомого камподея, сходного с представителями данного типа) - активные, нередко хищные, с развитыми придатками тела личинки. У них хорошо развиты три пары грудных ног, полностью расчлененных, длина их обычно превышает ширину тела. Сравнительно четко выделены грудной и брюшной отделы. Развиты антенны, характерны крупные верхние челюсти. Покровы обычно пигментированы. Этот тип личинок встречается у некоторых семейств отряда Жесткокрылые (жужелицы, стафилины, божьи коровки) и у всех Сетчатокрылых.
Червеобразные личинки - очень крупная и неоднородная группа. Тело у них удлиненное, слабо подразделено на грудной и брюшной отделы. Три пары грудных ног короткие, либо отсутствуют. По наличию ног и развитию головы выделяют три формы червеобразных личинок. Личинки с головой и ногами встречаются у насекомых ряда семейств отряда Жесткокрылые и имеют разнообразную внешность. Личинки Пластинчатоусых (жуки-хрущи и навозники) толстые, С-образно изогнуты; личинки жуков-листоедов заметно выпуклые сверху; личинки жуков-щелкунов и чернотелок (проволочники и ложнопроволочники) длинные, тонкие, цилиндрические, с уплотненными покровами. Личинки с головой и без ног встречаются также у некоторых представителей семейств отряда Жесткокрылые (златки, усачи, долгоносики), у большинства групп отряда Перепончатокрылые (наездники, осы, пчелы, муравьи) и у части видов отряда Двукрылые (большинство комаров). Личинки без ног и внешне выраженной головы есть только у ряда представителей в отряде Двукрылые (большинство мух). Головной отдел этих личинок втянут в переднегрудь, головная капсула недоразвита.
Гусеницеобразные личинки имеют удлиненное тело и большое количество коротких ног. В отличие от других личинок, у них помимо трех пар грудных ног развито несколько пар придаточных брюшных ног. Эти короткие нерасчлененные личиночные придатки у взрослых насекомых не проявляются. Выделяют две формы гусеницеобразных личинок. Настоящие гусеницы - личинки Чешуекрылых имеют от 2 до 5 пар брюшных ног. Гусеницы большинства бабочек имеют 5 пар ног, расположенных на 3 -6-м и 9-м сегментах брюшка. Однако у некоторых семейств число брюшных ног сокращается до 4 (хохлатки), 3 (совки-металловидки) и 2 пар (пяденицы). Ложно гусеницы - личинки некоторых семейств пилильщиков, представителей отряда Перепончатокрылые. У них развито большее количество брюшных ног -6-8 пар.
Строение и типы куколок
У насекомых с полным превращением личинка, окончив развитие, превращается в куколку. Личинки большинства насекомых окукливаются в почве, иногда открыто - на растениях или внутри них. Куколка - покоящаяся стадия, однако в ее теле идут интенсивные противоположные процессы: распад личиночных органов и тканей и формирование органов и тканей имаго. К концу развития куколка приобретает основные черты строения взрослого насекомого: развитые крылья, ноги, антенны, ротовой аппарат имаго, фасеточные глаза. Придатки компактно сложены и прижаты к телу. Тело нередко покрыто дополнительной защитной оболочкой. По этому признаку выделяют три основные типа куколок (рис. 4 ).
 Рис. 4. Типы куколок насекомых
Рис. 4. Типы куколок насекомых Открытые , или свободные, куколки не имеют оболочки; все придатки тела хорошо различимы, свободны. Наглядно проявляется сходство куколок с имаго. Этот тип куколок встречается у подавляющего большинства Жесткокрылых и Перепончатокрылых, а также у насекомых отряда Сетчатокрылые. Очень редко открытые куколки встречаются у Чешуекрылых и Двукрылых.
Покрытые куколки имеют тонкую кожистую полупрозрачную оболочку, через которую можно различить ноги, крылья, антенны и другие части тела. Оболочка образуется выделениями кожных желез личинки в момент окукливания. Этот тип куколок характерен для подавляющего большинства Чешуекрылых, части отряда Двукрылые, очень редко встречается у Жесткокрылых и Перепончатокрылых.
Скрытые куколки заключены в плотную непрозрачную оболочку - пупарий, через которую нельзя различить части тела. Эта оболочка представляет собой личиночную шкурку, которая не сбрасывается как обычно при окукливании, а только отслаивается от тела и служит защитным покровом. Скрытая куколка характерна для большей части Двукрылых (большинство мух).
Такое развитие называется аметаболией: вылупившаяся из яиц молодь выглядит так же, как взрослые насекомые (имаго), и отличается только меньшими размерами и недоразвитостью половых органов. Наблюдается оно у щетинохвосток и других примитивных насекомых. (21.05 Кб) Неполный метаморфоз . При неполном, или постепенном, метаморфозе жизненный цикл обычно включает три стадии яйцо, личинка (нимфа) и имаго. Похожа нимфа на имаго или нет, но взрослое насекомое появляется на свет непосредственно после очередной линьки этой активной возрастной стадии. Личинка может отличаться от имаго только недоразвитием крыльев и половых органов ротовые органы, рацион и среда обитания у них практически одинаковы. Такое развитие называется паврометаболией. Оно свойственно, например, прямокрылым и клопам. С другой стороны, взрослые стрекозы и поденки сухопутные летающие животные, а их нимфы (они называются наядами) живут в воде и снабжены особыми личиночными органами, сильно отличающими их от имаго. Такое развитие называется гемиметаболией. Полный метаморфоз . В этом случае, называемом голометаболией, в жизненном цикле четыре стадии яйцо (практически отсутствующее при яйцеживорождении), личинка, куколка и имаго. Личинка активно питается. Куколка внешне пассивна, но на этой стадии происходит радикальное преобразование личиночных структур в имагинальные. Взрослое насекомое никогда не похоже на личинку; куколка также почти всегда мало чем ее напоминает (самое близкое их сходство наблюдается у ряда мух). Во многих случаях запасов, накопленных личинкой, хватает имаго на весь остаток жизни. У высших мух окукливание происходит внутри личиночной оболочки, и образующаяся структура известна как пупарий. Иногда куколка окружена особым защитным чехлом коконом; она может находиться под землей или внутри кормового растения в специально устраиваемой куколочной камере. Большинство куколок неактивно, но многие из них перед самым выходом имаго с помощью движущихся при сокращениях тела шипов проделывают из куколочной камеры выход и иногда даже частично из нее высовываются. Обычно коконы очень прочны, однако у одного из концов в их стенке находится «слабое» кольцо. Вышедшее из куколки насекомое выделяет растворяющую это кольцо жидкость и открывает образующуюся в результате круглую дверцу. У молодых высших мух есть специальный заполненный жидкостью пузыревидный орган, расположенный над антеннами. Он используется для выталкивания наружу «дверцы» пупария, а также помогает животному прорыть выход на поверхность почвы. СЕНСОРНЫЕ ОРГАНЫ Слух . По крайней мере некоторые насекомые обладают очень тонким слухом, хотя большинство их не различает тонов, доступных для человеческого уха, их рецепторы настроены на гораздо более высокие частоты. У сверчков и кузнечиков органы слуха находятся на передних голенях, а у саранчовых у основания брюшка. Чем слышит большинство других насекомых, точно не известно, но звуковоспринимающие структуры расположены у них на различных частях тела. Возможно, многие виды вообще «глухи» в нашем понимании, но улавливают предупреждающие об опасности или присутствии добычи сигналы в форме вибраций поверхностей, на которых они сидят. Вероятно, такой способностью обладает большинство пещерных форм. Однако для многих насекомых слух крайне важен, поскольку сами они издают смысловые звуки, обеспечивающие внутривидовое общение, например привлекающие полового партнера. Так, самцы сверчков призывают самок «песнями», возникающими при т.н. стридуляции трении друг о друга передних крыльев, а саранчовые стридулируют, потирая о переднее крыло бедро задней ноги.Метаморфо́з (у насекомых), коренная перестройка внешнего и внутреннего строения насекомого в период постэмбрионального развития при переходе от личинки ко взрослому насекомому - имаго. В том случае, когда между личинкой и имаго возникает покоящаяся стадия, или куколка, перестройка организма личинки происходит именно в ней. В настоящее время различают следующие типы метаморфоза.
Анаморфоз характерен для отряда бессяжковых (Protura). Выходящая из яйца личинка, по сравнению со взрослым насекомым, имеет меньшее количество брюшных сегментов, недоразвитые ротовые органы и брюшные придатки. Недостающие сегменты образуются в процессе последовательных линек из особой зоны роста, расположенной на конце брюшка.
Протоморфоз наблюдается у представителей отрядов ногохвосток (Collembola), двухвосток (Diplura) и щетинохвосток (Thysanura). Выходящая из яйца личинка похожа на взрослое насекомое и имеет такое же количество сегментов, которые, однако, еще не разделены на грудные и брюшные. В процессе последовательных линек происходит их дифференциация, увеличение количества члеников усиков и церков, а также развитие половой системы. Отличительной чертой протоморфоза является линька во взрослом состоянии.
Гемиметаморфоз, или неполное превращение, характерен для представителей отрядов поденок (Ephemeroptera), стрекоз (Odonata), богомоловых (Mantoptera), термитов (Isoptera), веснянок (Plecoptera), эмбий (Embioptera), уховерток (Dermaptera), зораптер (Zoraptera), а также для большинства таракановых (Blattoptera), палочников (Phasmatoptera), прямокрылых (Orthoptera), сеноедов (Mallophaga), клопов (Hemiptera) и равнокрылых (Homoptera). Гемиметаморфоз характеризуется прохождением трех стадий развития - яйца, личинки и взрослого насекомого. Выходящая из яйца личинка похожа на взрослое насекомое, имеет тот же тип ротового аппарата и, чаще всего, обитает в тех же биотопах, что и имаго. Проходя определенное количество линек, она превращается во взрослое насекомое. За это время у нее развиваются крылья и формируется половая система. Для поденок характерно наличие двух крылатых стадий, разделенных линькой. Первая - субимаго - является неполовозрелой. Половая система развивается только после ее линьки в имаго. Из-за этого развитие поденок часто рассматривают как видоизмененный протоморфоз.
Неполное превращение имеет две модификации - гипо- и гиперморфоз. Гипоморфоз, или пониженное неполное превращение, наблюдается в отрядах тараканосверчков (Grylloblatidae), гемимерид (Hemimeridae), пухоедов (Mallophaga), вшей (Anoplura), а также у части таракановых, палочников, прямокрылых, клопов и сеноедов. Он характерен только для вторичнобескрылых насекомых (утративших крылья в процессе эволюции). Из-за отсутствия крыльев личинки очень сходны со взрослыми насекомыми, отличаясь незначительными деталями внешнего строения и отсутствием половой системы.
Гиперморфоз, или повышенное неполное превращение, наблюдается у трипсов (Thysanoptera), а также у алейродид (Homoptera: Aleyrodinea) и самцов кокцид (Homoptera: Coccinea). Его отличительной особенностью является покоящаяся личинка последнего возраста, рассматриваемая как аналог куколки. Некоторые исследователи рассматривают этот вариант как переходный к полному превращению.
Голометаморфоз, или полное превращение, характерен для
Неполное превращение, или гемиметаморфоз (греч. ημιμεταμόρφωση), в целом характеризуется прохождением лишь трех стадий - яйца, личинки и имаго. Личинки насекомых с неполным превращением внешне сходны со взрослыми особями и, подобно последним, имеют сложные глаза, такие же, как у взрослых, ротовые органы и в более старших возрастах - хорошо выраженные наружные зачатки крыльев. Помимо этого, у многих насекомых с неполным превращением личинки ведут сходный с имаго образ жизни и могут встречаться совместно с последними.
Вследствие большого морфологического и биологического сходства с имаго такие личинки называются имагообразными или нимфами (встречающееся иногда применение термина «нимфа» к личинкам лишь последних возрастов с развитыми крыловыми зачатками является некорректным). Нимфы веснянок и стрекоз претерпевают особую модификацию - они также похожи на имаго, но живут в воде и обладают провизорными, то есть специально личиночными, органами - жабрами и др. Таких личинок называют наядами.
В качестве подтипа гемиметаморфоза выделяют также его упрощение - гипоморфоз (лат. hypomorphosis), характерный для вторично бескрылых представителей насекомых с неполным превращением - вшей, пухоедов, некоторых тараканов, прямокрылых и др. При гипоморфозе нимфы внешне практически неотличимы от имаго. Прежде считалось даже, что эти насекомые развиваются без превращения.
Полное превращение, или голометаморфоз (греч. ολομεταμόρφωση), характеризуется прохождением от четырех до пяти стадий - яйца, личинки, куколки, имаго и иногда предкуколки.
Личинки насекомых с полным превращением, как правило, мало похожи на взрослых особей, поэтому называются истинными личинками. Действительно, трудно узнать в червеобразной личинке жука или гусенице бабочки взрослое крылатое насекомое. При этом личинка всегда лишена сложных глаз, наружных зачатков крыльев и часто имеет иной тип ротовых органов, нежели взрослые особи. Нередко сегменты тела истинной личинки однородны, то есть отсутствует резкое разделение тела на грудной и брюшной отделы. Помимо того, истинные личинки живут в иной среде, чем взрослые. В связи с этим большинство органов истинных личинок имеют временный или, как говорят, провизорный характер, выполняя функции чисто личиночной жизни. От этих органов у взрослых насекомых обычно не остается и следа. К числу провизорных органов личинок относятся: брюшные ноги, ротовой аппарат, паутинные железы и т. д.
Основная масса истинных личинок относится к трем группам:
Камподеовидные - внешне сходны с двухвостками, подвижные, обычно темноокрашенные личинки с плотными покровами и тремя парами грудных ног, хорошо обособленной прогнатической головой и часто со щетинками на конце тела (жужелицы, плавунцы, сетчатокрылые и др.);
Червеобразные - малоподвижные, светлоокрашенные личинки, лишенные брюшных и грудных ног или с короткими грудными ногами (двукрылые, многие перепончатокрылые, многие жуки);
Гусеницеобразные , или эруковидные, - умеренно подвижные личинки с тремя парами грудных ног и 2-8 парами брюшных (личинки скорпионниц, гусеницы, ложногусеницы).
Другая классификация истинных личинок основывается на тех стадиях развития эмбриона, при прохождении которых происходит вылупление из яйца:
Полиподные - то же, что гусеницеобразные личинки;
Олигоподные - камподеовидные и червеобразные личинки, имеющие грудные ноги;
Апоидные - безногие червеобразные личинки.
Несходство истинных личинок со взрослой фазой распространяется и на образ жизни - питание, место обитания и пр.
Закончив свой рост, истинная личинка последнего возраста прекращает питание, становится неподвижной, линяет в последний раз и превращается в куколку. Иногда куколке предшествует особая стадия, называемая предкуколкой. По сути это последний возраст истинной личинки, переход в который сопровождается утратой подвижности, сокращением размеров и началом процесса гистолиза задолго до линьки в куколку.
Анаморфоз (греч. αναμόρφωση) - одна из двух первичных форм метаморфоза насекомых, возможно, свойственная наиболее примитивному из отрядов насекомых - бессяжковым. При анаморфозе личинки внешне очень сходны со взрослыми, но имеют меньшее число брюшных сегментов. С развитием личинки происходит нарастание дополнительных сегментов на вершине брюшка, но полное их число достигается лишь во взрослой фазе. Этот тип превращения широко распространен у многоножек и других «низших» членистоногих. Следует заметить, что наличие настоящего анаморфоза у бессяжковых неоднократно подвергалось сомнению. Помимо увеличения с линьками числа брюшных сегментов метаморфоз бессяжковых интересен также наличием стадии предымаго.
Первичное превращение , или протоморфоз (греч. πρωτομορφωση), - второй первичный для насекомых тип метаморфоза, сохранившийся у ногохвосток, двухвосток, щетинохвосток и поденок. Характеризуется наличием двух взрослых стадий: субимаго и имаго. Личинка в целом сходна со взрослой стадией.
Гиперморфоз (греч. υπερμόρφωση) - модификация гемиметаморфоза, являющаяся в некотором роде преходной к полному превращению. Характерен для белокрылок, кокцид и трипсов. Его отличие от неполного превращения состоит в появлении в конце развития личинки стадии покоя, называемой иногда ложнокуколкой. Также нимфа этих насекомых (особенно у самцов кокцид) не настолько похожа на взрослое насекомое, как у насекомых с типичным гемиметаморфозом. Однако на стадии покоя при гиперморфозе всё же не происходит настолько сложной перестройки строения, как это происходит при полном превращении.
Физиология метаморфоза. Метаморфоз сопровождается, помимо внешних, также и внутренними изменениями. Важнейшую роль при метаморфозе играют гормоны.
При неполном превращении внутренние изменения протекают постепенно и при переходе во взрослую фазу не сопровождаются коренной перестройкой всей личиночной организации; многие органы личинок сохраняются у имаго. Даже зачатки яичников и семенников обнаруживаются уже у личинок первого возраста, развиваются постепенно и при переходе во взрослую стадию у одних видов превращаются во вполне зрелые половые железы, у других доразвиваются во взрослом состоянии.
Совсем иной характер внутренних изменений имеют насекомые с полным превращением. Их личинки резко отличаются от имаго, поэтому переход во взрослую стадию неизбежно требует коренной перестройки всей морфологической и биологической организации. Эта перестройка происходит в основном в стадии куколки и слагается из двух процессов - гистолиза и гистогенеза.
Сущность гистолиза состоит в уничтожении личиночных органов, сопровождаемым проникновением и внедрением в ткани гемоцитов. Содержимое куколки как бы возвращается к недифференцированному состоянию яйца. Гистолизу подвергаются все системы организма личинки, кроме нервной, половой, а также спинного сосуда.
В дальнейшем гистолиз сменяется гистогенезом, конечной целью которого является построение из образовавшейся жидкой массы новых, имагинальных органов. Важную роль при гистогенезе также играют имагинальные зачатки - группы клеток, из которых возникают те или иные ткани и органы.
Существеннейшую роль при метаморфозе играет эндокринная система. Рост и развитие насекомых регулируются нейросекреторными клетками головного мозга, кардиальными телами, прилежащими телами и переднегрудными железами. Мозговой гормон через аксоны мозга переносится к кардиальным телам, а оттуда - в кровь. Он стимулирует обмен веществ, а также переднегрудные железы, вырабатывающие гормон линьки экдизон. Прилежащие тела выделяют ювенильный гормон, который препятствует линьке во взрослую фазу и стимулирует рост и развитие личиночных органов. С возрастом рост тела личинки опережает рост ювенильных тел, и их роль постепенно затухает. Поступление ювенильного гормона в кровь прекращается и насекомое вступает в стадию имаго.
Результатом проводившихся экспериментов по пересадке ювенильных тел в личинку последнего возраста у прямокрылообразных и клопов стало получение дополнительных, ненормально крупных личинок. Так в условиях опыта удается увеличить число личиночных возрастов.
Фазовая изменчивость Особую форму метаморфоза, проходимую совместно с основным, представляет собой так называемая фазовая изменчивость, открытая первоначально у саранчовых, но затем обнаруженная и у остальных прямокрылых, а также у палочников, бабочек и других насекомых. Сущность её заключается в том, что при возникновении скученности особей изменяются морфологические и биолого-физиологические свойства вида - усиливается пигментация покровов (в основном из-за частичного отложения в них продуктов метаболизма), изменяются пропорции и форма частей тела, ускоряется развитие, меняется поведение. Возникает стадная фаза (лат. phasis gregaria). При рассеивании особей эти свойства утрачиваются и вид возвращается через переходную фазу (лат. phasis transiens) в свое исходное состояние - одиночную фазу (лат. phasis solitaria). Отличия между фазами столь велики, что незнание их природы порождало разделение одного вида на 2-3.
Функциональная роль стадий метаморфоза. Для насекомых характерно разделение функций не только в пространстве (дифференциация органов), но и во времени: в течение онтогенеза происходит не только смена функций отдельных органов, но меняются и основные функции организма как целого.
Для личиночных стадий насекомых характерна функция питания, накопления пластических и энергетических веществ, а для имаго - функция размножения. Функция расселения может выполняться как имагинальной, так и личиночными стадиями. Большинству насекомых, как несомненно прогрессивной группе, свойственно активное расселение, осуществляемое полётом. Поэтому весь облик имаго большей части крылатых насекомых - это облик расселяющейся стадии.
Иногда имагинальная стадия выполняет только функции видовой жизни - расселительную и размножения, утрачивая функцию питания. Однако выполнение функций расселения и размножения часто протекает в различные сроки; часто расселение происходит в те сроки, когда половые продукты еще не созрели. Так, самки многих совок совершают перелёты, когда их яичники еще не созрели. Более того, после созревания половых продуктов они отяжелевают настолько, что теряют способность даже к небольшим взлётам. Расселительную функцию иногда имеют и брачные полеты, особенно у общественных насекомых. Таким образом, у многих насекомых можно говорить о возрастном функциональном разделении стадии имаго на две «подстадии»: расселительную и репродуктивную.
Само собой разумеется, что между всеми рассмотренными типами специализации есть переходы, но они вопреки ожиданию не столь многочисленны.
Места обитания. Н асекомые обитают в подавляющем большинстве известных наземных биотопов, заняв такие негостеприимные экосистемы, как высокогорье, глубокие пещеры а также зарождающиеся экосистемы недавно образовавшихся островов вулканического происхождения. Известны и морские насекомые, относящиеся к особому семейству водомерок из отряда полужесткокрылых (кроме них, в прибрежных соленых водах редко поселяются другие, типично пресноводные клопы).
Избирательность стаций (местообитаний) является весьма существенным и характерным свойством насекомых. Каждый вид имеет свой набор стаций - в одних случаях разнообразный, в других ограниченный экологическими рамками и иногда сведенный к единственному типу местообитания. Однако часто вид не ограничивается заселение только лишь одной стации: проявляется закономерное изменение видами своих местообитаний. Смена стаций может быть зональной, вертикальной, сезонной и годичной.
Зональная смена стаций свойственна многим трансзональным видам (то есть видам, ареал которых пересекает несколько природных зон): при продвижении к северу ареала избираются более сухие, хорошо прогреваемые открытые местообитания, в то время как при продвижении к югу заселяются более увлажненные и затененные стации, часто с густым растительным покровом. Особой формой зональной смены стаций является зональная смена ярусов - переход в сухих частях ареала наземных видов к подземному образу жизни. Вертикальная смена стаций аналогична зональной, но характерна для горных условий. Если горная система подпирается аридными ландшафтами - степями или пустынями, а низинная часть ареала отодвинута от горной к северу, тогда с повышением уровня виды переходят в более увлажненные стации. При контакте низинной и горной частей ареала повышение вертикального уровня вызывает перемещение видов из более влажных стаций в менее увлажненные и более открытые.
Сезонная и годичная смены стаций протекают уже во времени являются следствием вынужденных миграций вида в результате изменений микроклимата, погодных условий и состояния растительного покрова. Первая характерна для областей с жарким летом и заключается в переходе в более увлажненные стации. Годичная смена стаций вызывается отклонением климатических показателей от средней нормы и приводит в сухие годы к перемещению ряда видов во влажные стации и, наоборот, во влажные годы - к перемещению в сухие стации.
Экологической основой смены стаций является строгое подчинение своему экологическому стандарту. В результате изменяется тип заселяемых стаций, что приводит в конечном итоге к изменению экологии вида, стимулирует внутривидовую дифференциацию и становится фактором эволюции. Нередки случаи, когда в разных зонах вид представлен разными подвидами или даже дифференцировался на два очень близких вида.
Географически увеличенная стация представляет собой ареал. Для многих насекомых известны сплошные ареалы, хотя в классе, как выясняется в последнее время, относительно высока доля видов-двойников, порой различающихся лишь по биохимическим особенностям. Также, как и ареалы других животных, ареал насекомых характеризуется принадлежностью к одной или нескольким зоогеографическим областям. Некоторые исследователи полагают, что можно создать единую подробную схему зоогеографического районирования, приемлемую для всех насекомых. Другие считают, что создание такой универсальной схемы невозможно, поскольку даже такая единая в таксономическом отношении группа, как насекомые, не представляет собой чего-то единого в отношении закономерностей географического распространения. Поэтому бессмысленно пытаться создать единую зоогеографическую карту, а следует разрабатывать карты для отдельных экологических групп насекомых. В частности, такая карта для реофильных амфибиотических насекомых (большинство поденок) совершенно несопоставима с картой для растительноядных наземных насекомых и существенно отличается от карты лимнофильных амфибиотических насекомых, как, например, стрекозы.
Для общей характеристики расположения ареалов самых разных насекомых обычно используют широко признанные названия крупных разделов суши: Палеарктика, Неарктика, Голарктика, Амфипацифический сектор, Эфиопская область, Ориентальная область, Арктогея, Неотропики и Австралийская область.
Размножение. Насекомые раздельнополы. Органы размножения самки обычно представлены парными яичниками и тянущимися по бокам яйцеводами, которые сливаются в один непарный проток, впадающий во влагалище. У самок имеются семяприемники и придаточные половые железы. У самцов имеются парные семенники, от которых отходят семяпроводы, тянущиеся по бокам тела. В нижней части семяпроводы расширяются, образуя семенные пузырьки, предназначенные для хранения спермы. Семяпроводы объединяются в общий семяизвергательный канал, открывающийся на способном увеличиваться или выдвигаться совокупительном органе. Придаточные железы секретируют семенную жидкость.
Роль звуков велика в жизни насекомых, хотя изучена слабо. Большой вклад в её изучение внесли биоакустики Е. К. Еськов, А. В. Попов, Р. Д. Жантиев.
Поскольку насекомые имеют малые размеры и не обладают единым развитым дыхательным трактом, в котором мог бы разместиться источник звука (гортань), они задействуют другие способы звукоизвлечения. Например, трение задней ноги (имеющей специальные бугорки) об надкрылья, как саранча, или надкрылий между собой (где левое обычно исполняет роль смычка), как у кузнечиков и сверчков. Звуки некоторых представителей последних могут разноситься до полутора километров. Цикады используют специальную звуковую мембрану, приводимую в колебание мышцами. Получающийся металлический звук резонирует в специальных полостях внутри тела, достигая большой громкости. Южно-американская цикада способна издавать звук, похожий на свист паровоза. Термиты при опасности в большом числе бьют головами о субстрат (материал термитника), создавая барабанный бой и оповещая таким образом других термитов.
Медоносные пчёлы издают звуки, заставляя вибрировать часть торакса путём частых мышечных сокращений. Звук усиливается крыловыми пластинами. В отличие от многих насекомых пчёлы способны издавать звуки разной высоты и тембров, что позволяет им передавать информацию посредством разных характеристик звука.
Роль в природе . Насекомые составляют около 80 % всех животных на Земле, по разным оценкам в современной фауне от 2 до 10 млн видов насекомых, из них пока известно чуть более 1 млн. Активно участвуя в круговороте веществ, насекомые играют глобальную планетарную роль в природе.
Более 80 % растений опыляются насекомыми, и можно с уверенностью сказать, что цветок - результат совместной эволюции растений и насекомых. Приспособления цветковых растений для привлечения насекомых разнообразны: пыльца, нектар, эфирные масла, аромат, форма и окраска цветка. Приспособления насекомых: сосущий хоботок бабочек, грызуще-лижущий хоботок пчелиных; особые пыльцесобирательные аппараты - у пчел и шмелей щеточка и корзинка на задних ногах, у пчел-мегахил - брюшная щеточка, многочисленные волоски на ногах и теле.
Огромную роль насекомые играют в почвообразовании. Такое участие связано не только с разрыхлением почвы и обогащением ее перегноем почвенными насекомыми и их личинками, но и с разложением растительных и животных остатков - опада растений, трупов и экскрементов животных, одновременно выполняется санитарная роль и круговорот веществ в природе.
Санитарную роль выполняют следующие виды насекомых: копрофаги - жуки-навозники, мухи-навозницы, коровницы; некрофаги - жуки-мертвоеды, могильщики, кожееды, мухи-мясоедки, падальницы; насекомые - разрушители отмерших растительных остатков: древесины, веток, листьев, хвои - жуки-сверлильщики, личинки усачей, златок, рогохвостов, комаров-долгоножек, муравьи-древоточцы, грибные комарики и т. д.; насекомые - санитары водоемов питаются взвешенными или осевшими на дно гниющими органическими веществами (детритом) - личинки комаров-дергунов, или звонцов, поденок, ручейников, очищают воду и служат биоиндикатором ее санитарного состояния.
Происхождение. Установление происхождения класса насекомых проблематично. Ключевая проблема состоит в отсутствии окаменелостей, по которым можно было бы установить филогенетические отношения насекомых.
Традиционно на основе морфологических сравнений (в особенности трахеальных дыхательных систем) ближайшими родственниками насекомых считали многоножек. В соответствии с этой точкой зрения обе группы объединяли в таксон «неполноусые» (Atelocerata).
Однако построение филогений на основе последовательностей генов и недавние морфологические исследования свидетельствуют, что насекомые ближе к ракообразным, чем к многоножкам. Сторонники этой филогенетической гипотезы объединили ракообразных и насекомых в единый таксон Pancrustacea. Данная гипотеза также проблематична. Ракообразные известны из отложений раннего Кембрия (около 511 млн лет), в то время как насекомые появляются в палеонтологической летописи только в раннем Девоне (около 410 млн лет). Таким образом, гипотеза о происхождении ракообразных и насекомых от единого общего предка порождает временной промежуток в ок. 100 млн лет, в течение которого насекомые должны были существовать. Отсутствие останков насекомых в палеонтологической летописи в течение указанного периода обычно объясняется тем, что они не сохранились до наших дней.
Новейшие морфологические сравнения и филогенетические реконструкции на основе геномных последовательностей указывают, что насекомые являются потомками ракообразных, а не сестринским таксоном. Это заключение хорошо согласуется с палеонтологическими данными. Однако морфологические и молекулярные данные не согласуются при определении ближайших родственников насекомых среди ракообразных: морфологические данные указывают на связь насекомых с высшими ракообразными, а молекулярные - с жаброногими.
В соответствии с последней гипотезой эволюционная ветвь насекомых отделилась от ракообразных в позднем Силуре - раннем Девоне. Эта оценка согласуется и с палеонтологическими данными, и с оценкой на основе молекулярных часов.
Литература
Догель В.А. Зоология беспозвоночных. – М.: Высшая школа, 1981. – С. 339 – 399.
Рис. 2. Расчлененный жук-олень.

Рис. 3. Голова насекомого.


Рис. 4. Сяжки насекомых. Рис. 5. Ротовой аппарат (грызущего типа) насекомых.



Рис. 6. Строение грудного сегмента и ноги. Рис. 7. Жилкование крыла.




