
Каламиты-полностью вымершие растения. Они впервые появляются в верхнем девоне, наибольшего разнообразия и развития достигают в каменноугольном периоде, в нижней перми это семейство даже было в отдельных районах главным углеобразующим. Исчезают каламиты полностью в юрском периоде.
Некоторые виды каламитов были крупными деревьями достигавшими 30 м. в высоту и 1 м. в диаметре. Внешним обликом походили на современные хвощи т.к. являлись их двоюродными предками. Стебли (стволы) были членистыми и ребристыми (стволы некоторых видов были гладкие и лишь наличие мутовчато расположенных веточных рубцов говорит о членистости ствола).У молодых каламитов в стволах присутствовала сердцевина которая с возрастом (по официальной версии) разрушалась и стволы становились полыми. Но мои находки говорят, что скорее всего главным фактором разрушения сердцевины стали насекомые которые избрали сердцевину каламитов местом инкубации своих личинок.
фото-17. Место выхода насекомого из сердцевины каламита.
местонахождение: "скала-2", средний карбон, свита С-2/3 смоляниновская, песчаник интервала известняка Н5(Н6) - угля h9.
Ветви также членистые, на каждом узле несли сильно варьировавшее от вида к виду количество узких иногда ланцетовидных со срединной жилкой листьев (Annularia) со сросшемся или нет основаниями. Выполнявшими роль фотосинтеза в отличии от листьев современных хвощей.

фото-7. Листья и ветви каламитов.
фиг.1,2,3 - листья и ветви Asterophillytes longifolius f. striata.
фиг.4 - листья Calamites multiramis.
Органы размножения (Palaeostachya)

фото-12. Органы плодоношения.
фиг.1,1a,2,2a - фрагменты слороносных колосков Palaeostahia sp. "скала-2", средний карбон, башкирский ярус, свита С-2/3 смоляниновская, интервал известняка Н5(Н6) - угля h9, песчаник.
фиг.3 - схема строения спороносного колоска. С.В.Мейен "Основы палеоботаники" стр.92 рис.22 фиг."в".
и см. (фото-11 фиг.2) имели вид колосков разной конструкции и размера.Росли каламиты на заболоченной почве и имели длинные горизонтальные корневища

фото-13. Корневище каламита.
от которых отходили вверх надземные вертикальные побеги.

фото-9. Подземные части каламитов.
фиг.1 - реконструкция подземной части ствола эукаламита."Основы палеонтологии" под редакцией Орлова т.14
фиг.1а - подземная часть ствола Calamites (Calamitina sp.)."скала-1", средний карбон, башкирский ярус, свита С-2/3 смоляниновская, интервал известняка Н5(Н6)- угля h9, песчаник.
фиг.2 - реконструкция подземной части ствола стилокаламита. "Основы палеонтологии". под редакцией Орлова т.14
фиг.2а - подзкмная часть ствола стилокаламита. "скала-1", средний карбон, башкирский ярус, свита С-2/3 смоляниновская, интервал известняка Н5(Н6) - угля h9, песчаник.
фиг.3 - подзкмная часть ствола Calamites carinatus."скала-2", средний карбон, башкирский ярус, свита С-2/3 смоляниновская, интервал известняка Н5(Н6) - угля h9, песчаник.

фото-18. Место оветвления побега от корневища.
местокахождения "Балка лесная" средний карбон, сита С-2/4, кровля угольного пласта i-3/1в
Имеющиеся в коллекциях остатки стволов каламитов представляют собой внутренние ядра. На наружной поверхности этих ядер наблюдается более или менее отчётливые бороздки (расположенные на "рёбрах" хорошо видны на фото-17) , маркирующие направление бывших сосудов (проводящих пучков). По особенностям строения пучка в области узла порядок делится на три рода:
1.Род астерокаламитовые (Asterocalamites или более современное Archaeocalamites)

фото-1. Аrchaeocalamites sp. (al. Asterocalamites). В данном случае пoявляются признаки перехода к мезокаламитам.
к нему принадлежат более примитивные растения порядка, жившие от верхнего девона до нижнего карбона (моя находка двух экземпляров говорит что дотянули до низов среднего карбона, местонахождения приуроченные к свите С-2/3). У архаеокаламитов проводящие пучки пересекают и там соединяются слабо выраженными комиссурами. Листья были длиной до 5 см. узкие и много кратно дихотомически (раздваивались) ветвились.
2.Род мезокаламит (Mesocalamites).

фото-2. Mesocalamites ramifer.
фиг.1 - узел.
фиг.2 - ветвь (заросток)?
фиг.3,4 - сердцевинный отлив ствола.
фиг.5 - схема прохождения через узел каламита проводящих сосудов.C.В.Мейен "Основы палеоботаники" стр.90 рис.21 фиг."в".
Мезокаламиты были крупными деревьями о чём свидетельствует остаток ветки (см. фото-3)

фото-3. Ветвь мезокаламита.
Принадлежащие сюда каламиты представляют особенный интерес, так как по строению ствола они занимают промежуточное положение между Archaeocalamites и Calamites , причём это положение соответствует их стратиграфическому положению (верхи нижнего карбона - низы среднего карбона).Строение ствола этого рода характерно тем, что ход сосудистых (проводящих) пучков в очень варьировал. Местами сосуды проходили через узел чередуясь как у истинного каламита (см. фото-4 фиг.2)а местами как у архаеокаламитов (см. фото-1).
3.Род Каламит (Calamites).Который в свою очередь делится на две большие условные группы:
а).Эукаламиты

фото-4. Эукаламиты. Условная группа включающая себя множество видов. Особенностью данной группы было значительное превышение длины между узлия (расстояние между пережимами ствола) над его толщиной. И так называемое "правильное" ветвление.Эталонным представителм этой группы был Calamites carinatus (фиг.1) (на каждом узле противопоставлено распологались 2 крупных веточных рубца). Но у другого рода Calamitina перед узлом с ветвями длина между узлия уменьшалась (фиг.5). Иногда, при консолидации (уплотнении) осадка стволы каламитов лопались вдоль оси и в образовавшуюся щель выдавливался заполнявший внутреннюю полость ил. В результате образовывался своеобразный "киль" (фиг.7).
фиг.1 - реконструкция внешнего вида.Н.А.Комарницкий и др. "Систематика растений" стр.287 рис.146 фиг.2а.
фиг.2 - проводящие сосуды.
фиг.3 - плавающие корни Radicites sp.
фиг.4 - корневище.
фиг.5,6,7,8 - слепки внутренней полости стволов.
Местонахождения: старый каменный карьер (фиг.2), скала-1 (фиг.4,5,

фото-5. Минерализованный (интускрустация) фрагмент ствола Эукаламита.
фиг."а,б,в,г,е" - разные проэкции фрагмента.
фиг."д" - на сколе хрошо видны проводящие сосуды.

фото-14. Веточные рубцы эукаламитов.
C под-родом Calamitina

фото-6. Представитель эукаламитов подрод Calamitina goepperti.
фиг.1 - реконструкция внешнего вида. М.Хирмер (Hirmer. 1927).
фиг.2 - проэкции сердцевинного отлива (внутреннего ядра).
Б). Стилокаламиты

фото-8. Стилокаламиты (фиг.4) многочисленная условная группа. Один из ярких представителей этой группы Сalamites Sachsei (фиг.1,2). Особенностью этого вида было то что в отличии от С.carinatus длина между узлия значительно (в 3-5 раз) меньше ширины. Количество ветвей колебалось у разных видов от 3 до 30 шт. (всегда кратно 3) сидевших на мощном колоннообразном стволе через каждые 4-6 между узлий. Фрагменты стволов без веточных рубцов относят к C.sukowi (фиг.3). Хотя на мой взгляд это очередной пример разных названий одного вида.
фиг.1,1a - сердцевинный отлив ствола.
фиг.2,2a - ствол С.Sachsei.
фиг.3 - ствол С.sukowi.
фиг.4 - реконструкция С.Sachsei.М.Хирмер (Hirmer.1927)
В ископаемом виде отпечатки коры и сама кора встречаются крайне редко.
Местонахождения: скала-1 (фиг.1,1а)-средний карбон свита С-2/3(смоляниновская); балка "лесная" (фиг.2,2а,3)- средний карбон свита С-2/4.

фото-10. Заросток Calamites multiramis.
Одной из самых больших проблем палеонтологии является разрозненность остатков организмов будь то животное или растение.И находка частей организма в прикреплении или хотя бы в близкой ассоциации большая удача! И вот мне на местонахождении "Балка лесная" удалось найти в ассоциации практически все вегетативные части Calamites Sachsei

фото-11 (фото-7 в публикации МЕСТОНАХОЖДЕНИЕ "БАЛКА ЛЕСНАЯ") . Стерильный (фиг.1) , фертильный (фиг.2) побеги и ствол Calamites sachsei.
В пермском периоде род Calamites вымирает но не задолго до этого его пермские потомки

фото-16. Пермские паракаламиты.
фиг.1 - Paracalamitina sp. (P1 , Новый Кувак)
фиг.2 - Paracalamites decoratus. (P1. Красно-Усольск)
по не понятным причинам (возможно так повлияли изменения климата и изменения среды обитания) возвращаются в схеме проводящих сосудов к примитивным формам своих предков.

фото-15. Сравнение строения проводящих сосудов каламитов.
фиг.1 - Archaeocalamites sp.("скала-1", средний карбон, свита C-2/3 смоляниновская,песчаник,интервал известняка H5(H6)- угля h9.
фиг.2 - Mesocalamites sp.("скала-1", средний карбон, свита C-2/3 смоляниновская.
фиг.3 - Calamites (Calamitina) Sachsei.("старый каменный карьер", средний карбон, свита C-2/3 смоляниновская, песчаник, интервал известняка Н5(Н6) - угля h9.
фиг.4 - Parocalamites decoratus (P1, Красноусольск).
Растения, входящие в отдел хвощевидных (Equisetophyta ), отличаются от других споровых побегами, составленными отдельными члениками (междоузлиями) и узлами с мутовчато расположенными листьями. Характерной чертой хвощевидных является также наличие у них спорангиофоров – особых структур, несущих спорангии. Мутовки спорангиофоров образуют на стебле спороносные зоны, чередующиеся с обычными вегетативными листьями, или сидят на концах осей, образуя стробилы.
Ископаемые остатки хвощевидных известны из девонских отложений. К концу этого периода хвощевидные уже прошли длительный путь эволюции, а расцвет этой группы растений пришелся на каменноугольное время. В палеозое хвощевидные отличались исключительным разнообразием: среди них встречались травы, кустарники, лианы и деревья, достигавшие 20 м высоты. Вместе с гигантскими плауновидными эти формы образовывали каменноугольные леса.
В то время, помимо представителей класса хвощевых (Equisetopsida ), на Земле произрастали и хвощевидные растения другого класса – клинолистовых (Sphenophyllopsida )1 . Клинолисты (род Sphenophyllum ) образовывали густые заросли в лесах каменноугольного периода (рис. 1). Побеги некоторых растений этой группы достигали в длину нескольких метров при очень небольшом диаметре (0,1–0,5 см) и были либо полегающими, либо лазающими, о чем свидетельствуют прицепки, обнаруженные на стеблях одного из видов. К узлам молодых участков стебля прикреплялись мутовки из 6–9 клиновидных листьев.

1,2 – клинолист клинолистный (Sphenophyllum cuneifolium) (1 – общий вид части растения, 2 – листья разных узлов стебля); 3–5 – стробилы представителей порядка клинолистовых
(3 – сфенофиллостахис водяной (Sphenophyllostachys aquensis ); 4–5 – сфенофиллостахис Досона (S. dawsonii ), (4 – продольный срез части стробила, 5 – поперечный срез стробила в области междоузлия; сп – спорангий, сс – стерильный сегмент спорангиофора, вс – веточка спороносного сегмента спорангиофора)
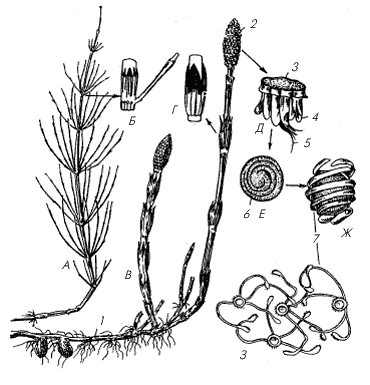
Древние представители класса хвощевых – каламиты (Calamites ) – в каменноугольном периоде в значительной мере составляли древесный ярус заболоченных тропических лесов. По внешнему виду каламиты напоминали современные хвощи, только гигантских размеров (рис. 2). Стволы каламитов достигали 10–20 м высоты и 0,5 м в диаметре за счет способности к вторичному утолщению (слой вторичной древесины в их стеблях достигал 12 см).
1–3 – каламиты (Calamites) с разными типами ветвления; 4 – листья каламитов; 5 – поперечный срез стробила каламита; 6 – часть стробила, вид сбоку: слева – с поверхности, справа – в продольном разрезе (б – брактея, пп – проводящий пучок, сф – спорангиофор, сп – спорангий)
Листья, основные фотосинтезирующие органы каламитов, длиной от 2 до 7 см, были расположены в узлах стебля мутовками по 3–70 штук. Стробилы, длиной около 12 и шириной около 4 см, состояли из чередующихся мутовок стерильных листьев и спорангиофоров. Возможно, все каламиты были разноспоровыми растениями, т.е. формировали мужские и женские споры, а на образовывавшихся из них гаметофитах (заростках), соответственно, развивались либо только мужские, либо только женские половые клетки. У одного из каламитов (каламокарпон замечательный – Calamocarpon insignis ) найден мегастробил, в спорангии которого развивалась всего одна мегаспора. Она прорастала прямо внутри спорангия, образуя женский гаметофит. Такая крайняя степень редукции числа мегаспор в спорангии отмечена и у древовидных плауновидных палеозоя 2 .
В отличие от большинства древовидных плауновидных и класса клиноситовых каламиты пережили каменноугольный период и были обильно представлены и в пермское время. Но к началу мезозойской эры каламитовые, достигшие высокой степени специализации, вымерли под воздействием резкого изменения климата. Их менее специализированные сородичи из класса хвощовых на некоторое время пережили климатические перемены и в первой половине мезозоя в некоторых областях Земли даже образовали почти чистые сообщества по берегам водоемов. Однако и они постепенно почти все вымерли, так что в наше время весь отдел хвощевидных представлен одним родом хвощ (Equisetum ), относящимся к классу хвощовых и включающим около 20 видов.
Латинское название Equisetum впервые употребил Плиний Старший, имея в виду, по-видимому, сходство ветвистых побегов хвоща с хвостом лошади (от лат. equius – лошадь и saeta, seta – щетина, жесткие волосы). В русском названии «хвощ» отражено сходство взрослого растения с хвостом или с пучком волос. В некоторых местностях России хвощи с мутовчато расположенными побегами называют «сосенка», «елка», а по наличию у некоторых видов бурых спороносных побегов, похожих на пест, – «пестиками» или «толкачиками».

1 – участки горизонтального и вертикального корневища с клубнями у хвоща полевого; 2 – то же у хвоща болотного; 3 – схема поперечного среза клубня хвоща полевого; 4 – схема поперечного среза корня хвоща полевого (вп – воздушная полость, пп – проводящий пучок, пт – покровная ткань)
Все современные хвощи – многолетние корневищные травы с мутовками бурых редуцированных листьев, утративших хлорофилл (не следует путать с листьями мутовки зеленых веточек в узлах стеблей). Корневища у всех хвощей глубоко залегающие, сильно разветвленные, в их узлах часто закладываются почки. Почки на корневищах нередко образуют короткие утолщенные междоузлия (называемые клубнями), складывающиеся в короткие цепочки, богатые крахмалом. От узлов корневищ отходят многочисленные придаточные корни двух типов: растущие вниз, часто утолщенные, и направленные вверх или в стороны (рис. 3). Стебли прямостоячие или восходящие, простые или мутовчато-ветвистые, цилиндрические или с продольными ребрами, состоят из узлов и полых междоузлий. В узлах побегов находятся листовые влагалища, по верхнему краю которых расположены зубцевидные листовые пластины (рис. 4, В , Г ; 6). Листья хвощей срастаются основаниями в кольцо, сидящее на узле и прикрывающее растущую зону междоузлий, богатую сахаром. Верхушки листьев имеют форму узких зубцов. В почке эти зубцы сращены и образуют колпачок, который при росте стебля разрывается на верхушке. В дальнейшем при росте стебля в длину и толщину, листовые пластинки отходят друг от друга, но разрыв происходит иногда не по всем линиям срастания, поэтому число зубцов по краю влагалища может не соответствовать действительному числу листьев. Форма влагалища, цвет, форма и время жизни зубцов различны у разных видов, и эти признаки используются при определении хвощей.
А – вегетативный побег; Б – спороносный побег; В, Г – листовые влагалища; Д – спорангиофор со спорангиями; Е–З – споры (1 – корневище с клубнями, 2 – стробил; 3 – щиток; 4 – спорангий; 5 – ножка; 6 – наружная оболочка споры, 7 – элатеры)
В основании листовых влагалищ хвощей закладываются почки, которые позже прорывают влагалища и образуют боковые побеги. Для возобновления растения особенно важны почки на тех узлах стеблей, которые находятся под землей, вблизи ее поверхности. Рядом с почкой закладывается корень.
По характеру надземных побегов хвощи можно разбить на две группы. У одних видов все надземные побеги однотипны. Они очень жесткие, обычно вечнозеленые (т.е. живут более одного сезона и в умеренной зоне перезимовывают) и развивают верхушечные стробилы.
У видов второй группы побеги бывают двух типов: одни – спороносные – буроватые или зеленые, а другие – вегетативные, зеленые. Побеги обоих типов появляются весной над поверхностью почвы, они нежнее по своей консистенции, чем побеги первой группы, и к зиме отмирают. Развитие вегетативных и спороносных побегов у видов второй группы идет по-разному. У хвоща полевого спороносные побеги появляются первыми ранней весной (рис. 4, В ). Они буровато-розовые, неветвистые, обычно толще быстро зеленеющих вегетативных побегов и после спороношения большей частью отмирают. У хвощей лесного и лугового спороносные побеги с хорошо развитыми стробилами появляются ранней весной одновременно с вегетативными. И те и другие вначале бледно-розовые, а затем зеленеют. После спороношения и отсыхания стробила спороносные побеги функционируют подобно вегетативным. Наконец, у таких видов, как хвощ болотный, хвощ приречный, хвощ боготский (E. bogotense – он обитает в тропиках Центральной и Южной Америки), спороносные побеги в момент их появления над поверхностью земли с трудом можно отличить от вегетативных – у них такая же зеленая окраска и сходные размеры.

Рис. 5. Формы листовых мутовок (а) и поперечные срезы побегов (б) хвощей:
1 – хвощ полевой; 2 – хвощ луговой; 3 – хвощ лесной; 4 – хвощ болотный; 5 – хвощ зимующий (а – листовое кольцо; б – поперечный разрез стебля)
Жесткость побегов хвощей объясняется тем, что в состав оболочки клеток эпидермы и, в меньшей степени, других тканей побега наряду с целлюлозой входит кремнезем. Частицы кремнезема располагаются в клеточной оболочке между тяжами целлюлозы, а на поверхности клеток эпидермы образуют сплошной и гомогенный, очень прочный слой с мелкими бугорками, приуроченными к выростам оболочки (рис. 5). Слой из кремнезема покрыт снаружи тонкой кутикулой с восковым налетом и играет не только механическую, но и защитную роль, так как благодаря ему хвощи практически не подвергаются нападению различных растительноядных моллюсков и насекомых.
Стробилы или спороносные колоски хвощей состоят из спорангиофоров, размещенных на общей оси. Они имеют вид шестиугольных щитков с ножками (рис. 4, Д ). На внутренней стороне такого щитка размещены 4–6 спорангиев. Когда находящиеся в них споры созревают, ось колоска немного удлиняется, и щитки спорангиофоров отходят друг от друга. Стенки созревших спорангиев лопаются, открывается продольная щель, и споры высыпаются. Лентовидные придатки (элатеры), образующиеся из наружной оболочки споры, способствуют сцеплению распространяющихся спор друг с другом (рис. 4, Ж , З ).
Споры могут прорастать только на затененной и влажной почве, где из них вырастает пластинчатый или ветвистый заросток. Заростки (гаметофиты) современных хвощей – очень маленькие, размером в несколько миллиметров, недолговечные зеленые растеньица.
Современные хвощи в отличие от вымерших каламитов – равноспоровые растения, на их заростках образуются органы (архегонии и антеридии), продуцирующие как женские (яйцеклетки), так и мужские (многожгутиковые сперматозоиды) половые клетки.
Сперматозоиды хвощей подвижны, поэтому оплодотворение происходит в воде – во время дождя или выпадения росы. Из зиготы развивается зародыш, а из него – без периода покоя – проросток, который через несколько месяцев превращается во взрослую особь нового бесполого поколения – спорофит (рис. 6).

Рис. 6. Чередование поколений в жизненном цикле хвощей (Equisetum arvense):
а – гаметофит (заросток); б – спорофит; в – спорангофор; г – спорангии; д – щиток спорангиофора; е – элатеры; ж – клубеньки
В природе хвощи обычно встречаются в виде клонов (групп растений, возникших путем вегетативного размножения), которые нередко занимают участки площадью в несколько десятков и даже сотен квадратных метров.
Хвощи отличаются крайним полиморфизмом. Например, у хвоща полевого, недавно поселившегося на хорошо освещенном участке с нарушенным растительным покровом, стебли обычно низкие, распростертые. Но позже, когда разовьется окружающая растительность, от более зрелых корневищ того же клона могут отходить мощные прямостоячие стебли с ветвистыми побегами. При приближении к северным границам ареала этого вида спороносные побеги, буроватые у растений умеренной зоны, сменяются зеленеющими, а потом – в тундре – и ветвистыми зелеными. Очень изменчивы бывают побеги и у других видов. Эти модификации, возникающие под влиянием многообразных факторов внешней среды (часто даже в пределах одного клона), наряду с многочисленными гибридами и наследуемыми уродствами, являются одной из причин разногласий между ботаниками в оценке объема рода.
Хвощи довольно широко распространены на Земле, особенно в Северном полушарии. Но в Австралии, Новой Зеландии и Тропической Африке аборигенных видов хвощей нет.
Встречаются хвощи в различных растительных сообществах, но обязательно с достаточным или избыточным увлажнением. По берегам водоемов, в сырых лесах, в некоторых типах низинных болот хвощи нередко доминируют в травянистом покрове.
Хвощи часто захватывают территории с нарушенным растительным покровом, образуя при этом чистые или почти чистые заросли в тех местах, где другие растения не могут жить, например, из-за обилия воды. Раз поселившись на какой-нибудь территории, хвощи, благодаря наличию глубоко залегающих корневищ, масса которых превышает массу надземных частей растения в несколько раз, успешно противостоят таким неблагоприятным воздействиям внешней среды, как засухи, лесные пожары и т.п., и успешно конкурируют с другими растениями, зачастую становясь широко распространенными и трудно искореняемыми сорняками пастбищ и полей.
Хвощи, растущие в условиях избыточного увлажнения, обладают признаками гигрофитов – у них слабо развита водопроводящая система, но хорошо представлена воздухоносная ткань. Одновременно эти растения могут нести и признаки ксерофитов – погруженные под поверхность эпидермы замыкающие клетки устьиц, закрытые устьица на старых участках стебля, сильно развитая механическая ткань, фотосинтезирующие стебли, редуцированные листья. Соотношение тех или иных признаков строения варьирует у разных видов.
В умеренной зоне России чаще всего можно встретить следующие виды хвощей.

Рис. 7. Хвощ луговой (Equisetum pratense Ehrh.)авт. рис. Шикина А.В.
Хвощ луговой (E.pratense, рис. 7) – многолетнее растение с удлиненным подземным черным корневищем без клубней. Спороносные побеги бледно-зеленые, простые, с воронковидными крупными влагалищами и свободными светло-коричневыми зубцами, с перепончатой каймой. Колосок длиной 0,8–2 см, цилиндрический. Спороношение в июне-июле. После спороношения, в начале лета, из спороносных побегов развиваются простые зеленые ветви, и они становятся похожими на вегетативные. Вегетативные побеги хвоща лугового имеют высоту 10–30 см и диаметр около 2 мм, они прямостоячие, также бледно-зеленые, внутри с большой центральной полостью и многими мелкими периферическими (рис. 5), с 8–14 шероховатыми ребрами, с простыми неветвящимися боковыми трехгранными веточками длиной 5–15 см, расположенными горизонтально или дугообразно книзу. Зубцы листовых мутовок широколанцетные, на ветвях яйцевидные, острые, в количестве 10–15 шт.
Продолжение следует
Общая характеристика хвощевидных
Одно из названий этих очень древних растений - членистостебельные - очень точно характеризует особенности их строения. Побеги хвощевидных состоят из четко оформленных стеблей с междоузлиями и узлами с мутовчато расположенными на них мелкими чешуевидными листьями.
Вот этими стеблями ныне живущие хвощи и ископаемые виды отличаются от всех остальных бессемейных высших растений и немного похожи на водоросли (харовые), голосеменные (хвойник) и даже цветковые (казуарину).
Хвощевидные, как будто собранные из деталей конструктора, «суставчатые» растения, заметно выделяются на фоне травостоя в лесу, на лугу или в поле.
Латинское название хвощевидных Equisetophyta обязано древнеримскому естествоиспытателю Плинию Старшему. Он нашел некоторое сходство ветвистых побегов хвоща с хвостом лошади (equius - лошадь, saeta, seta - щетина).
Во многих европейских языках название хвощей переводится тоже как «конский хвост». Например, польское название звучит как konsciogon, английское - horsetail и французское - queue - de cheval.
В русском названии хвощевидные тоже проявляются отзвуки однокоренных слов - «хвост», «хвощ». За отдельными видами хвощей с зелеными, мутовчато расположенными листьями в некоторых регионах нашей страны закрепились названия елка, сосенка. А пестиками, толкачиками люди называют весеннее поколение хвощей - буровато-желтые спороносные побеги.
Древние предки хвощевидных - каламиты - были огромными деревьями, высотой до 20 м, с мощными ветвистыми и ребристыми стволами и были похожи на гигантские хвощи (см. рис. учебника, с. 59). В каменноугольный период каламиты вместе с древовидными плаунами и папоротниками образовывали целые ярусы заболоченных тропических лесов.
Между стволами порхали гигантские стрекозы, другие насекомые, которые в то время были единственными властителями воздушной стихии; по земле неуклюже передвигались древние земноводные - первые завоеватели земной суши.
Современные представители хвощевидных - многолетние корневищные травы. Среди них есть и небольшие, карликовые виды и много довольно высоких растений. У хвоща камышкового высота стеблей 5–15 см, а их диаметр 0,5–1 мм. Стебли другого вида - хвоща многощетинкового достигают 9 м, а диаметр равняется 0,5–2 см. Такие высокие и тонкие стебли с недостаточно развитой механической тканью не могут самостоятельно пребывать в прямостоячем положении, поэтому растут, опираясь на другие растения.
У хвощевидных вырастают стебли двух видов: зеленые вегетативные и спороносные бесхлорофилльные, которые быстро отмирают. И зеленые, и бесхлорофилльные побеги имеют продольные борозды и расчленены, как и корневища, на узлы и полые междоузлия.
От узлов отходят мутовки ветвей и много мелких, часто чешуевидных листьев, которые срастаются в трубочки, охватывающие стебли. Чешуевидные листочки у спороносных побегов бесхлорофилльные. Спорангии формируются на верхушках стробилов.
Из спор вырастает очень маленький зеленый гаметофит, размером всего несколько миллиметров. На поверхности такой крошечной зеленой пластинки только при наличии влаги осуществляется оплодотворение, после чего из объединенной клетки - зиготы без периода покоя развивается новое поколение - спорофит хвоща.
Распространены хвощи очень широко, почти по всему земному шару, кроме Австралии, произрастая в самых разных растительных сообществах и зонах, но обязательно рядом с водой. Это могут быть участки с достаточным содержанием влаги в почве либо с относительно неглубоким залеганием грунтовых вод.
Еще одной характерной особенностью роста и развития хвощевидных является их обильное разрастание на больших площадях в результате вегетативного размножения даже одного-единственного экземпляра хвоща. От корневищ такого «отца» или «матери» всех будущих вегетативных потомков отрастают все новые и новые побеги, которые в свою очередь развивают новые корневища с новыми побегами. Так что большие заросли хвощей часто являются клонами одного растения и имеют одни и те же наследственные свойства.
В формировании флоры, растительного покрова нашей планеты , в настоящее время хвощи, как и плауновидные, не имеют большого значения. Роль хвощей может быть даже отрицательной. Быстро размножаясь в местах с нарушенным растительным покровом, хвощевидные становятся трудно искореняемыми сорняками пастбищ и полей. Хвощи быстро отрастают даже после тщательной прополки на полях и огородах, потому что на их хорошо развитых корневищах имеются специальные образования в виде подземных клубней, которые называют земляными орешками или земляными шишками. Благодаря таким особенностям хвощи переносят и лесные пожары.
В большинстве своем хвощевидные несъедобны для животных. Более того, кормление лошадей сеном с большим количеством хвощей может быть причиной заболевания, которое в народе называется «шатун» или «пьяная болезнь». Это заболевание проявляется в параличе ног и в нарушении движения лошадей.
Но кабаны и олени осенью и зимой иногда подкрепляются хвощами (летом они их не поедают), видимо, активность ядовитых веществ в увядших и подмерзших хвощах значительно снижается.
| <<< Назад
|
Вперед >>>
|



