Комплементарным называется такой вид взаимодействия неаллельных генов, при котором действие гена из одной аллельной пары дополняется действием гена из другой аллельной пары, в результате чего формируется качественно новый признак.
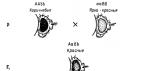
Классический пример такого взаимодействия – наследование формы гребня у кур. Встречаются следующие формы гребня: листовидный – результат взаимодействия двух рецессивных неаллельных генов аabb; ореховидный – результат взаимодействия двух доминантных неаллельных генов A-B-; розовидный и гороховидный – c генотипами A-bb и aaB- , соответственно.
Другой пример – наследование окраски шерсти у мышей. Окраска бывает серая, белая и черная, а пигмент только один – черный. В основе формирования той или иной окраски шерсти лежит взаимодействие двух пар неаллельных генов:
A – ген, определяющий синтез пигмента;
a – ген, не определяющий синтез пигмента;
B – ген, определяющий неравномерное распределение пигмента;
b – ген, определяющий равномерное распределение пигмента.
Примеры комплементарного взаимодействия у человека: ретинобластома и нефробластома кодируются двумя парами неаллельных генов.
Возможные варианты расщепления в F 2 при комплементарном взаимодействии: 9:3:4; 9:3:3:1; 9:7.
Эпистаз
Эпистаз - такой вид взаимодействия неаллельных генов, при котором действие гена из одной аллельной пары подавляется действием гена из другой аллельной пары.
Различают две формы эпистаза – доминантный и рецессивный. При доминантном эпистазе в качестве гена-подавителя (супрессора) выступает доминантный ген, при рецессивном эпистазе – рецессивный ген.
Пример доминантного эпистаза – наследование окраски оперения у кур. Взаимодействуют две пары неаллельных генов:
С – ген, определяющий окраску оперения (обычно пеструю),
с – ген, не определяющий окраску оперения,
I – ген, подавляющий окраску,
i – ген, не подавляющий окраску.
Варианты расщепления в F 2: 12:3:1, 13:3.
У человека примером доминантного эпистаза являются ферментопатии (энзимопатии) – заболевания, в основе которых лежит недостаточная выработка того или иного фермента.
Пример рецессивного эпистаза – так называемый «бомбейский феномен»: в семье у родителей, где мать имела группу крови О, а отец – группу крови А, родились две дочери, из которых одна имела группу крови АВ. Ученые предположили, что у матери в генотипе был ген I B , однако его действие было подавлено двумя рецессивными эпистатическими генами dd.
Полимерия
Полимерия - такой вид взаимодействия неаллельных генов, при котором несколько неаллельных генов определяют один и тот же признак, усиливая его проявление. Это явление противоположно плейотропии. По типу полимерии обычно наследуются количественные признаки, чем и обусловлено большое разнообразие их проявления в природе.
Например, окраска зерен у пшеницы определяется двумя парами неаллельных генов:
A 1
a 1 – ген, не определяющий красную окраску;
A 2 – ген, определяющий красную окраску;
a 2 – ген, не определяющий красную окраску.
A 1 A 1 A 2 A 2 – генотип растений с красной окраской зерен;
a 1 a 1 a 2 a 2 - генотип растений с белой окраской зерен.
Расщепление в F 2: 15:1 или 1:4:6:4:1.
У человека по типу полимерии наследуются такие признаки, как рост, цвет волос, цвет кожи, величина артериального давления, умственные способности.
Эффект положения
Эффект положения – вид взаимодействия неаллельных генов, обусловленный местом положения гена в генотипе.
Пример – наследование белка Rh- фактора (резус-фактора). У 85% европейцев резус-фактор имеется (Rh+ ), у 15% – его нет (Rh- ). Определяется резус-фактор тремя доминантными генами (С, D, E), расположенными в хромосоме рядом друг с другом.
Два человека с одинаковым генотипом CcDDEe будут иметь разные фенотипы в зависимости от варианта расположения аллельных генов в паре гомологичных хромосом: в варианте А – много антигена Е, но мало антигена С; в варианте В – мало антигена Е, но много антигена С.
Вариант А Вариант В
1. Схема взаимодействия между генами
2. Типы доминирования
3. Комплементарные гены
4. Генотипы мышей, фенотипическое проявление результата скрещивания
1. Развитие любых признаков у организмов является следствием сложных взаимодействий между генами, точнее, между продуктами их деятельности - белками-ферментами. Эти взаимодействия могут быть представлены в виде следующей схемы:
взаимодействие генов одной аллельной пары:
Неполное доминирование;
Доминирование;
Сверхдоминирование;
Кодоминирование;
Взаимодействие генов различных аллельных пар:
Комплементарное действие;
Эпистаз;
Полимерия.
Доминированиепроявляется в тех случаях, когда одна аллель гена полностью скрывает присутствие другой аллели. Однако, по-видимому, чаще всего присутствие рецессивной аллели как-то сказывается и обычно приходится встречаться с различной степенью неполного доминирования. Очевидно, зуо объясняется тем, что доминантная аллель отвечает за активную форму белка-фермента, а рецессивные аллели часто детерминируют те же белки-ферменты, но со сниженной ферментативной активностью. Это явление и реализуется у гетерозиготных форм в виде неполного доминирования.
Сверхдоминирование заключается в том, что у доминантного аллеля в гетерозиготном состоянии иногда отмечается более сильное проявление, чем в гомозиготном состоянии.
Кодоминирование - проявление в гетерозиготном состоянии признаков, детерминируемых обеими аллелями. Например, каждый из аллельных генов кодирует определенный белок, и у гетерозиготного организма синтезируются они оба. В таких случаях путем биохимического исследования можно установить гетерозиготность без проведения анализирующего скрещивания.
Этот метод нашел распространение в медико-генетических консультациях для выявления гетерозиготных носителей генов, обусловливающих болезни обмена. По типу кодоминирования у человека наследуются группы крови.
3. Сложные отношения возникают между неаллельными парами генов (комплементарное действие, эпистаз, полимерия и т. д.).
Комплементарными называются взаимодополняющие гены. Их примером может служить скрещивание двух рас душистого горошка, имеющих белый ивет :
Гибриды первого поколения оказались не белыми, а красно-фиолетовыми;
Во втором поколении обнаружилось неожиданное расщепление в соотношении 9:7.
Генетический анализ показал, что окраска цветов душистого горошка зависит от двух комплементарных генов. Каждый из них доминантен, но в отсутствие другого гена своего действия не проявляет. Генотип одной расы горошка с белыми цветами был Aabb, другой - ааВВ. При скрещивании их гибриды имели генотип АаВЬ, и тогда окраска проявилась.
Во втором поколении все растения с доминантными аллелями обоих генов оказываются окрашенными, но растения, имеющие лишь доминантный аллель одного из генов, как и имеющие только рецессивные аллели этих генов, - однотипными, бесцветными.
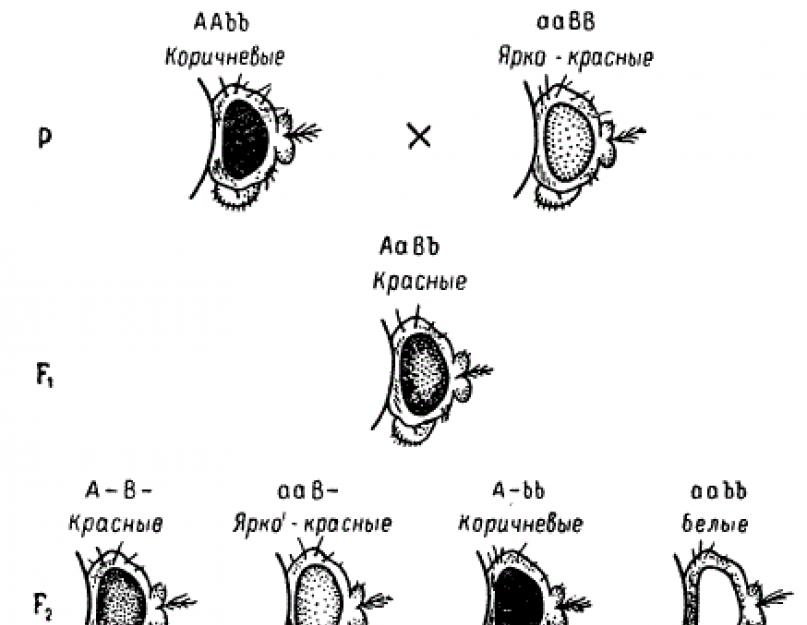
4. Своеобразный результат обнаружен при скрещивании черных и белых мышей. Все особи первого поколения были серыми. А во втором поколении расщепление произошло в соотношении 9:3:4. Выяснилось, что окраска шерсти у мышей также контролируется двумя комплементарными генами. Но в отличие от предыдущего примера один из генов (А) имеет собственное фенотипическое появление, второй же (В) реализуется фенотипически лишь в присутствии первого.
Доминантный аллель А необходим для синтеза пигмента, в отсутствие его (аа) пигмент не развивается и животные оказываются альбиносами. Доминантный аллель В обеспечивает отложение пигмента в волосе в форме черных колец, вследствие чего волосы приобретают серую окраску. Если доминантный аллель В отсутствует в зиготе, т. е. по этому гену животное имеет генотип bb, то при наличии доминантного аллеля А пигмент в волосах откладывается равномерно и они приобретают черную окраску.
Альбиносы, взятые в опыт, были гомозиготами по рецессивному гену окраски и по доминантному гену зонального распределения пигментов (ааВВ). Черные мыши были гомозиготными по доминантному гену окраски и рецессивному гену зонального распределения пигментов (Aabb). Мыши в F1 имели генетическую конституцию АаВЬ и приобрели серую краску.
Для уяснения отмеченной закономерности генотипы мышей из F2 рекомендуется выписать на решетку и убедиться, что для особей, обладающих двумя доминантными генами (ААВВ), частота встречаемости равна 9/16 (они все имеют серую окраску), для особей, имеющих доминантный аллель первого гена и рецессивный второго (Aabb) - 3/16 (черные). Наконец, соотношение потомков, получивших только рецессивные аллели первого гена и доминантные второго (ааВВ), как и особей, несущих рецессивные аллели обоих генов (aabb), составит 3/16 + 1/16, т. е. 4/16 (белые). Аналогичные случаи наследования встречаются у многих видов животных и растений.
К комплементарным , или дополнительным , генам относят такие гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состояниях (А-В-) обусловливают развитие нового признака.
Действие же каждого гена в отдельности (А-bb или ааВ-) воспроизводит признак лишь одного из скрещиваемых родителей.
Впервые такого рода взаимодействие было обнаружено у душистого горошка (Lathyrus odoratus). При скрещивании двух рас этого растения с белыми цветками у гибрида F 1 цветки оказались пурпурными. При самоопылении растений F 1 в F 2 наблюдалось расщепление по окраске цветков в отношении, близком к 9:7. Один фенотипический класс (9 / 16) имел такую же окраску цветков, как и у растений первого поколения, а второй (7 / 16) - белую окраску, такую же, как у родительских растений.
Чтобы выяснить, укладывается ли это расщепление в схему дигибридного менделевского расщепления, представим себе, что у каждой исходной расы душистого горошка имеется в гомозиготном состоянии лишь по одной из доминантных аллелей (AAbb и ааВВ), которые при взаимодействии определяют развитие окраски. Поскольку у гибрида первого поколения присутствуют доминантные аллели обоих генов (АаВb), цветки гибридных растений F 1 будут окрашенными. Во втором поколении происходит расщепление в отношении 9 / 16 А-В-: 3 / 16 А-bb: 3 / 16 ааВ-: 1 / 16 aabb. Каждый из генов в отдельности не может обусловить развитие окраски, так как выработка антоциановых пигментов осуществляется лишь при наличии доминантных аллелей обоих генов. Поэтому растения с генотипами А-bb, ааВ- и aabb имеют белые цветки и во втором поколении наблюдается расщепление по фенотипу в отношении 9: 7. Анализирующим скрещиванием и анализом в F 3 можно точно подтвердить данное выше объяснение.
Приведем еще несколько примеров, иллюстрирующих действие комплементарных генов у растений и животных.
У земляники развитие «усов», т. е. вегетативных самоукореняющихся побегов, определяется доминантной аллелью, а «безусость» - рецессивной. Но существуют такие формы безусой земляники, которые при скрещивании друг с другом дают гибрид F 1 с сильно выраженным признаком «усатости». Исследованиями Т. С. Фадеевой было показано, что в потомстве такого гибрида в F 2 получается расщепление, близкое к отношению 9: 7, а именно: из 752 растений F 2 419 оказались с усами, 333 - без усов. Это соответствует теоретически ожидаемому расщеплению: 752 X 9 / 16 = 423 и 752 X 7 / 16 = 329.
У белого клевера имеются формы с высоким и низким содержанием цианида. Цианиды, как известно, блокируют дыхательный фермент, но повышают активность папаина (растительной протеазы), катепсина и других ферментов. Высокое содержание цианида в белом клевере связано с усиленным вегетативным ростом без снижения его кормовых качеств. При скрещивании растений с высоким и низким содержанием цианида в F 1 доминирует первое свойство, а в F 2 наблюдается расщепление, близкое к отношению 3: 1.
Эти результаты указывают на то, что в данном случае альтернативные признаки определяются одной парой аллелей. Но иногда при скрещивании двух растений клевера с низким содержанием цианида гибриды F 1 характеризуются высоким его содержанием, а в F 2 расщепление оказывается близким к отношению: 9 / 16 - с высоким содержанием цианида и 7 / 16 - с низким. Так же, как у душистого горошка, в данном случае имеет место обычное дигибридное расщепление, в котором 9 / 16 потомков обладают двумя доминантными генами А-В-, а 7 / 16 относятся к трем остальным фенотипически неотличимым классам: 3 / l 6 A-bb + 3 / 16 ааВ + 1 / 16 aabb = 7 / 16 . Доминантные аллели разных генов в отдельности не увеличивают содержание цианида по сравнению с тем низким уровнем, который характерен для растения, гомозиготного по рецессивным аллелям обоих генов, но при совместном действии доминантных аллелей обоих генов содержание цианида повышается.
Подобное явление можно показать на примере кукурузы. При скрещивании некоторых форм кукурузы с белыми зернами в F 1 зерна в початках оказываются пурпурными. В F 2 происходит расщепление на 9 / 16 пурпурных (А-В-) и 7 / 16 белых (ааВ-, А-bb и aabb).
До сих пор мы рассматривали примеры комплементарного взаимодействия доминантных генов, при котором каждый из генов в отдельности не обладал способностью вызывать развитие признака. Последний развивался лишь в результате взаимодействия доминантных аллелей двух генов. В силу этого в F 2 обнаруживались только два фенотипических класса в соотношении 9:7. Известны, однако, случаи, когда один или оба комплементарных гена характеризуются самостоятельным проявлением. В соответствии с этим меняется и характер расщепления в F 2 .
Рассмотрим наследование трех типов окраски шерсти у мышей: дикой, или рыжевато-серой (агути), черной и белой. Окраска дикого типа зависит от наличия гена, определяющего развитие окраски, и от гена, обусловливающего распределение пигмента по длине волоса. Каждый волос у мышей агути имеет по длине кольцо желтого пигмента, а в основании и на конце волоска - черный пигмент. Такое зонарное распределение пигментов и создает окраску агути, свойственную диким грызунам (белка, кролик, морская свинка и др.). У черных мышей отсутствует зонарное распределение пигмента - волосы до всей длине окрашены равномерно. Белые мыши с красной радужной оболочкой глаз, так называемые альбиносы, лишены пигмента.
Надо сказать, что альбинизм встречается у животных почти всех классов - млекопитающих, птиц, амфибий и др. Встречается альбинизм и у человека. Так, например, иногда у родителей-негров рождаются дети альбиносы, т. е. с белой кожей и белыми волосами, но с чертами лица негритянского типа. Известны случаи, когда в семье негров рождаются двойни (разнояйцевые), и один из детей оказывается альбиносом. Такой ребенок имеет рецессивную аллель гена альбинизма в гомозиготном состоянии.
Окраска шерсти у мышей типа агути доминирует над черной, и над белой. При скрещивании черных мышей с белыми с белыми, все гибриды F 1 оказываются агути, а в F 2 наблюдается расщепление в отношении 9 / 16 агути: 3 / 16 черных: 4 / 16 белых.
Взятые в скрещивание мыши-альбиносы являются, очевидно, гомозиготными по рецессивной аллели гена окраски и доминантной аллели гена попарного распределения пигмента (ааВB), а черные мыши - гомозиготными по доминантной аллели гена окраски и рецессивной аллели гена распределения пигмента в волоске (AAbb). У гибридов F 1 (АаВb) вследствие взаимодействия доминантных аллелей обоих генов развивается окраска типа агути. Такая же окраска характерна и для 9 / 16 особей в F 2 с генотипом А-В-. Черными в F. оказываются мыши, имеющие генотип А-bb, а белыми - все остальные - (ааВ- и aabb) в силу отсутствия у них гена А, определяющего образование пигмента. Ген В в отсутствие гена А не имеет собственного проявления.
Подобные примеры наследования известны и у растений (лук, кукуруза и др.). У лука скрещивание формы, имеющей неокрашенную (белую) луковицу, с формой, имеющей желтую луковицу, дает в F 1 растения с красными луковицами, а в F 2 появляются растении с красными (9 / 16), желтыми (3 / 16) и белыми (4 / 16) луковицами. В этом случае опять-таки одна из доминантных аллелей двух генов способна действовать самостоятельно (определяет желтую окраску луковицы), а другой ген проявляется лишь в присутствии комплементарного гена.
Известны и такие случаи, когда каждый из двух комплементарных генов способен проявлять свое действие самостоятельно. Один таких примеров мы уже рассматривали при анализе наследования формы гребня у кур. Каждая из доминантных аллелей генов обусловливала развитие гребня определенной формы (гороховидной или розовидной), а взаимодействие этих генов определяло развитие новой формы гребня ореховидной. В данном примере каждый из комплементарных доминантных генов характеризуется собственным специфическим эффектом, а взаимодействие между ними приводит к новообразованию, к новому выражению признака.
Ряд подобных примеров наследования известен и у других животных и растений. Так, у дрозофилы рецессивная аллель гена scarlet в гомозиготном состоянии определяет ярко-красную окраску глаз, а рецессивная аллель другого гена - brown (также в гомозиготном состоянии) определяет коричневую окраску глаз. При скрещивании гибриды F 1 оказываются красноглазыми (дикого типа), рели же оба эти рецессивных гена находятся в гомозиготном состоянии, то такая особь оказывается белоглазой. Если скрестить красноглазых мух F 1 друг с другом, то во втором поколении по признаку окраски глаз будет наблюдаться расщепление на 4 фенотипических класса в отношении 9 / 16 красных: 3 / 16 ярко-красных: 3 / 16 коричневых: 1 / 16 белых. Такое поведение признаков в наследовании также говорит о расщеплении по двум комплементарным генам с самостоятельным действием.

Если генотип мух с коричневыми глазами условно обозначить ААbb, с ярко-красными - ааВВ, а генотип красноглазых гибридов F 1 - АаВb и белоглазых мух - aabb, то фенотипические радикалы полученных в F 2 классов могут быть представлены как А-В- (9 / 16), ааВ- (3 / 16), А-bb(3 / 16) и aabb(1 / 16).
Природа взаимодействия генов в этом случае более ясна, чем в случае наследования формы гребней у кур. Нормальная красная окраска глаз у мух обеспечивается в основном тремя видами пигментов красным, коричневым и желтым. В гомозиготном состоянии рецессивный ген а блокирует образование коричневого пигмента, вследствие чего развиваются ярко-красные глаза, а другой рецессивный ген b в гомозиготном состоянии блокирует одновременно образование красного и желтого пигментов, и поэтому развиваются коричневые глаза. В F 1 объединяются доминантные аллели этих генов, поэтому образуются все пигменты, дающие в совокупности красную окраску глаз. Новый класс белоглазых мух, появляющихся в F 2 , очевидно, является результатом одновременного блокирования синтеза всех трех пигментов.
Подобные примеры можно привести и на растительных объектах. Известно, что окраска плодов у томатов обусловливается каротиновыми пигментами (ликопины и бета-каротин), имеющими огромное значение в синтезе витаминов. Анализ наследования окраски плодов у томатов показывает, что красная окраска плодов определяется взаимодействием комплементарных доминантных генов R и Т, оранжевые плоды образуются на растениях с генотипом R-tt, желтые - на растениях с генотипом rrТ-, промежуточные, желтооранжевые - на растениях rrtt. Здесь также расщепление в F 2 соответствует генетической формуле дигибридного скрещивания 9:3:3: 1. При этом установлено, что красные и оранжевые плоды содержат наибольшее количество каротинов, а желтые - наименьшее. Двойной рецессив содержит промежуточное количество каротинов в плоде. Качественные различия в наборе каротинов соответствуют определенным различиям в генотипе.
Во всех разобранных примерах мы имели дело с комплементарным взаимодействием доминантных и рецессивных неаллельных генов. Взаимодействие доминантных генов обусловливало развитие ореховидного гребня у кур, красной окраски глаз у дрозофилы, красной окраски плодов у томатов. Взаимодействие рецессивных аллелей этих генов приводило к развитию пластинчатого, или ликвидного, гребня у кур, белых глаз у дрозофилы, желто-оранжевой окраски плодов у томатов.
Следует отметить, что в ряде случаев комплементарные гены, способные к самостоятельному проявлению, при отсутствии дополнительного гена могут давать каждый в отдельности сходный фенотипический эффект. Характер расщепления дигибрида в F 2 при этом также изменяется. Так, у тыквы (Cucurbita реро) имеются сорта с разной формой плода: сферической, дисковидной и удлиненной. Сферическая форма плода является рецессивной по отношению к дисковидной.

От скрещивания растений с плодами сферической формы, но имеющими разное происхождение, возникают гибридные растения, дающие плоды только дисковидной формы. В потомстве этих растений в F 2 появляются три фенотипических класса в отношении 9/16 дисковидными плодами, 6/16 - со сферическими и 1 / 16 - с удлиненными. Зная закономерности дигибридного расщепления при взаимодействии генов, нетрудно понять, что и здесь имеет место взаимодействие двух генов, влияющих на развитие формы плода, каждый из доминантных комплементарных генов обусловливает развитие плодов сферической формы, а их взаимодействие приводит к образованию дисковидных плодов. Взаимодействие рецессивных аллелей этих генов определяет развитие плодов удлиненной формы.
Рассматривая примеры комплементарного действия генов, мы убеждаемся, что такое взаимодействие генов приводит к развитию признаков, свойственных диким предкам данных видов (серая окраска грызунов, дисковидная форма у тыквы и т. д.). Некоторые авторы рассматривают это явление как пример атавизма. Эти представления основываются на предположении, что в процессе эволюции животных и растений доминантные гены, действующие комплементарным образом, изменились, мутировали в рецессивное состояние (А → а, В → b, С → с и т. д.).
У диких предков домашних животных и растений доминантные гены комплементарного действия поддерживались естественным отбором вместе в одном генотипе (например, серая окраска грызунов, дисковидная форма плода у тыквы, красная окраска глаз у дрозофилы и др.). При одомашнивании и проведении селекции с помощью скрещиваний и искусственного отбора комплементарные гены как бы разобщились. Генотип АаВb разлагался селекционерами на генотипы ААЬЬ и ааВВ. Поэтому при скрещивании таких организмов иногда наблюдается как бы возврат к признакам диких предков.
Мы остановились более подробно на комплементарном действии генов потому, что этот тип взаимодействия иллюстрирует один из путей возникновения комбинативной изменчивости и имеет отношение к широко используемому явлению гибридной мощности - гетерозису.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
В том случае, когда признак появляется только при сочетании двух доминантных аллелей разных генов (например, А и В) их взаимодействие называют комплементарностью , а сами гены комплементарными (дополняющими друг друга). При этом каждый из взаимодействующих неаллельных генов в отсутствии другого не обеспечивает формирования признака. Комплементарное взаимодействие двух генов можно обозначить в виде формулы: Аа Вв
Известным примером комплементарного взаимодействия неаллельных генов является наследование окраски цветков душистого горошка (Lathyrus odoratus) при скрещивании двух родительских форм с белыми цветками ААвв и ааВВ. В потомстве F1 (АаВв), а также в F2 (фенотипический класс А-В-) будет появляться новая окраска – пурпурная.
При этом в F2 соотношение классов с окрашенными цветками (А-В-) и классов с неокрашенными цветками (А- bb; ааВ- и ааbb) будет соответствовать формуле 9:7 . Основными пигментами, определяющими окраску цветков душистого горошка, являются антоцианы.
Аналогичным примером является образование коричневого пигмента у шелкопрядов. Известно, что синтез пигмента ксантомматина (пигмент оммохромового ряда) осуществляется из триптофана.У шелкопряда известны рецессивные мутации двух неаллельных генов, которые, будучи в гомозиготном состоянии (генотипы ааВВ или ААbb) делают насекомых неокрашенными, поскольку мутации в любом из генов А или В блокирует синтез пигмента, а промежуточные соединения L-кинуренин и 3-оксикинуренин не имеют окраски. У гибридов первого поколения (АаВb) синтез пигмента восстанавливается в результате комплементарного взаимодействия генов А и В. В F2 наблюдается расщепление 9:7. По такому же принципу наследуется содержание цианида у растений клевера. У земляники развитие «усов», т. е. вегетативных самоукореняющихся побегов, определяется доминантным аллелем, а «безусость» – рецессивным. Но существуют такие формы безусой земляники, которые при скрещивании друг с другом дают гибриды F1 с сильно выраженным признаком «усатости». Было показано, что в потомстве такого гибрида в F2 наблюдается расщепление, близкое к отношению 9:7. Это наиболее простые примеры комплементарного взаимодействия неаллельных генов, когда действие каждого из них в отдельности вообще не проявляется. Признак развивается лишь в результате взаимодействия доминантных аллелей двух неаллельных генов. В силу этого в F2 обнаруживается только два фенотипических класса в соотношении 9:7. Известны, однако, случаи, когда один или оба комплементарных гена характеризуются самостоятельным проявлением . В соответствии с этим меняется и характер расщепления в F2. Пример комплементарного действия генов при наследовании формы плода у тыквы (Cucurbita pepo). В связи с тем, что генотипы ААвв и ааВВ фенотипически не различимы, то они в сумме дают цифру 6. Дисковидная форма возникает в результате взаимодействия двух доминантных генов (А и В), а удлиненная форма плода – как следствие сочетания их рецессивных аллелей. Рис. 33. Наследование формы плода у тыквы при комплементарном взаимодействии двух генов (расщепление 9: 6: 1)

ааВВ ААвв сферической формы формы F1: АаВв дисковидной формы F2: 9 А-В- : 3 ааВ- : 3 А-вв: аавв дисковидная сферическая удлиненная. Расщепление 9: 3: 4.
В том случае, когда фенотип одного из родителей (например, имеющего генотип ааВ-) совпадает с фенотипом рецессивной гомозиготы (ааbb), расщепление в F2 будет 9: 3: 4.
Наследование формы гребня у кур. В данном примере каждый из комплементарных доминантных генов характеризуется собственным специфическим эффектом, а взаимодействие между ними приводит к новообразованию, к новому выражению признака. Расщепление в F2 по фенотипу при этом полностью соответствует менделевскому отношению 9:3:3:1, ибо каждый из четырех классов (А-В-, А-bb, ааВ-, aabb) имеет свой особый фенотип. Откуда появляется дикий тип при скрещивании мутантных форм? Значит признаки комплементируют между собой (дополняют друг друга). К комплементарным, или дополнительным генам относят такие гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состояниях (А-В-) обусловливают развитие нового признака.
Действие же каждого гена в отдельности (А-bb или ааВ-) воспроизводит признак лишь одного из скрещиваемых родителей.
При скрещивании люцерны, имеющей красные цветки, с люцерной желтоцветковой в F x все растения оказываются с зелеными цветками. В F 2 получается: 9/16 с зелеными цветками, 3/16- с красными, 3/16 - с желтыми, 1/16 - с белыми цветками.
В этом скрещивании обнаружилась четкая картина дигибридного расщепления. Отсюда следует, что расщепление идет по двум парам генов. Но в данном случае доминантные гены К (красной окраски) и Ж (желтой окраски), действуя вместе (К + Ж), вызывают зеленую окраску цветков, т. е. зеленая окраска получается в результате взаимодополняющего (комплементарного) действия генов К и Ж. При наличии только одного доминантного гена К получаются красные цветки, одного гена Ж - желтые, только рецессивных генов (ккжж) - белые.
Комплементарное, взаимодополняющее действие генов встречается и у животных. Так, при скрещивании породы кур, имеющих розовидную форму гребня, с породой кур с гороховидным гребнем все потомство в F имеет ореховидный гребень. В F 2 получается 9/16 кур с ореховидным, 3/16 с розовидным, 3/16 с гороховидным и 1/16 с простым листовидным гребнем. В этом случае доминантные гены Р - розовидной, Г - гороховидной формы гребня, действуя вместе, дают новый признак - ореховидный гребень. Один ген Р вызывает розовидный, а один ген Г - гороховидный гребень. При наличии только рецессивных генов (рргг) образуется листовидный гребень.
Яркий пример комплементарного действия генов - скрещивание сортов душистого горошка, каждый из которых имеет белую окраску цветков. Их гибриды F] имеют яркоокрашенные красные цветки. В F 2 получается 9/16 растений с красными и 7/16 - с белыми цветками.
В этом примере один сорт горошка с белыми цветками имеет доминантный ген А , другой сорт горошка с белыми цветками имеет ген В. Ни ген А , ни ген В поодиночке не вызывают окраски цветков. При совместном же действии этих генов (А + В) у гибридов цветки ярко окрашиваются в красный цвет.
В F 2 у растений, имеющих оба гена (А + В ), цветки красные. Таких растений 9/16. У растений, имеющих лишь один из этих генов (либо А, либо В) и не имеющих их вовсе (аЪ ), цветки остаются белыми; таких растений оказывается 7/16.
Такие же проявления взаимодополняющего действия генов имеются у животных. Известны белые породы кур, дающие при скрещивании яр- коокрашенное потомство. В F 2 получается 9/16 окрашенных и 7/16 белых кур. Очевидно, в данном случае имеется также два гена. Ни один из них по отдельности не вызывает окраски оперения. Только при совместном их действии происходит окрашивание.
Интересная форма взаимодополняющего действия генов обнаруживается у тыквы. При скрещивании двух сортов тыкв, имеющих шаровидную форму плодов, получаются гибриды с дисковидными плодами. При разведении этих гибридов в следующем поколении появляется 9/16 растений с дисковидными плодами, 6/16 с шаровидными и 1/16 с удлиненными (рис. 2.1).
Рис. 2.1.
Результаты скрещивания двух сортов тыкв (рис. 2.1) объясняются следующим образом. Один сорт с шаровидными плодами имеет ген А, другой сорт с шаровидными же плодами - ген В. При их скрещивании у гибрида появляются гены А и В и получаются растения с дисковидными плодами. При их размножении в F 2 получается 9/16 растений, имеющих гены АВ (с дисковидными плодами), 3/16 имеют ген Л, но у них отсутствует ген В (с плодами шаровидной формы), 3/16 имеют ген В , но у них отсутствует ген А (также плоды шаровидной формы), 1/16 несут гены aabb (плоды удлиненной формы).
Наследование окраски шерсти при скрещивании черных и коричневых мышей можно изучить в ходе следующей работы.
У мышей окраска шерсти обусловлена многими взаимодействующими генами. Для масти агути (дикого типа) характерно кольцо желтого пигмента на каждом черном волоске. Образование его обусловлено геном А у который относится к серии множественных аллелей. Аллель а определяет отсутствие желтых колец на волосках. Мыши с генотипом аа имеют черную шерсть. Ген А оказывается доминантным относительно аллеля а , поэтому у животных с генотипом Аа окраска шерсти дикого типа. Над аллелями А и а доминирует ген этой же серии А у, который в гетерозиготном состоянии определяет желтую окраску, а в гомозиготном - летальность зародышей.
Окраску шерсти обусловливает и другой несцепленный ген - 6, который в гомозиготе вызывает образование коричневого пигмента. Он взаимодействует с генами A-а Если генотип имеет ген b (в гомозиготе) и ген А , то образуются желтые кольца на черных волосках и шерсть приобретает пестро-коричневую окраску.
Рецессивные гены а и b в гомозиготном состоянии (aabb) обусловливают новый признак - окраску шерсти шоколадного цвета. Следовательно, каждый фенотип обусловлен соответствующими генотипами: агути - А_В _., черные - ааВ_, коричневые - A bb , «шоколадные» - aabb.

При скрещивании черных и коричневых мышей схема скрещивания:
В первом поколении Fi все мыши серого цвета. При скрещивании мышей первого поколения между собой получаем:
|
ААВВ |
ААВЬ |
АаВВ |
АаВЬ |
||
|
ААВЬ |
ААЬЬ |
АаВЬ |
|||
|
Коричневые |
Коричневые |
||||
|
АаВВ |
АаВЬ |
ааВВ |
ааВЬ |
||
|
АаВЬ |
ааВЬ |
||||
|
Коричневые |
Шоколадные |
||||
В /*2 образуется четыре фенотипических класса (агути, коричневые, черные, «шоколадные».) В результате комплементарного взаимодействия генов а и b появляется новый признак - шоколадная окраска.
Теоретически числовое соотношение между классами составляет 9А_В_ : 3A bb : ЪааВ_ : 1 aabb.
Гены, которые видоизменяют действие других основных генов, сами по себе не определяя развитие признаков, называются генами-модификаторами.
Наследование окраски шерсти при тригибридном скрещивании белых и окрашенных мышей можно изучить в ходе следующей работы.

Окраска шерсти у мышей обусловлена многими генами. Из предыдущей работы известно взаимодействие генов А-а, В-b. Ген с обусловливает альбинизм у мышей. Г омозигота по этому гену (сс) лишена всякой окраски, независимо от наличия окрашивающих генов. Следовательно, генотипы альбиносов имеют разные гены окраски шерсти. Третья пара генов - Ос - не сцеплена с первыми, поэтому расщепляется независимо от них.
При скрещивании мышей серой окраски получаем:
|
ААВВСС |
ААВВСс |
АаВВСС |
ААВЬСС |
ААВЬСс |
АаВВСс |
АаВЬСС |
АаВЬСс |
|
|
ААВВСс |
ААВВсс |
АаВВСс |
ААВЬСС |
ААВЬсс |
АаВВсс |
АаВЬСс |
АаВЬсс |
|
|
АаВВСС |
АаВВСс |
ааВВСС |
АаВЬСС |
АаВЬСс |
ааВВСс |
ааВЬСС |
ааВЬСс |
|
|
ААВЬСС |
ААВЬСс |
АаВЬСС |
АаЬЬСС Коричневые |
ААЬЬСс Коричневые |
АаВЬСс |
АаЬЬСС Коричневые |
АаЬЬСс Коричневые |
|
|
ААВЬСс |
ААВЬсс |
АаВЬСс |
АаЬЬСс Коричневые |
ААЬЬсс |
АаВЬсс |
АаЬЬСс Коричневые |
||
|
АаВВСс |
АаВВсс |
ааВВСс |
АаВЬСс |
АаВЬсс |
ааВВсс |
ааВЬСс |
ааВЬсс |
|
|
АаВЬСС |
АаВЬСс |
ааВЬСС |
АаЬЬСС Коричневые |
АаЬЬСс Коричневые |
ааВЬСс |
ааЬЬСС Шоколадные |
ааЬЬСс Шоколадные |
|
|
АаВЬСс |
АаВЬсс |
ааВЬСс |
АаЬЬСс Коричневые |
ааВЬСс |
ааЬЬСс Шоколадные |
В результате скрещивания тригетерозиготных серых мышей получили: 27 АВ_С_ - серых;
- 9 А_ЬЬС_ - коричневых;
- 9 ааВ_С_ - черных;
- 3 ааЬЬС_ - шоколадных;
- 16 А_В_сс; ааВ сс; AJbbcc ; aabbcc - белых.
Задача 1, У душистого горошка гены Си? порознь вызывают белую окраску цветков, пурпурная же окраска получается только при наличии в генотипе обоих этих факторов. Растения с генотипом ссрр имеют белую окраску цветков. Какова будет окраска цветков в потомстве от скрещивания СсРр х ссРР?
В данной задаче наследование окраски обусловлено комплементарным взаимодействием двух генов.
Записываем кратко условие задачи:
Для определения фенотипов потомков сначала запишем гаметы родителей. Гетерозиготное материнское растение производит 4 типа гамет: СР, Ср , сР, ср , а отцовское гомозиготное растение - гаметы одного типа - сР. При слиянии женских гамет с мужскими образуется четыре типа генотипов:

Из схемы видно, что генотипы СсРР и СсРр содержат доминантные гены С и Р, которые обусловливают пурпурную окраску цветков. В остальных двух генотипах нет доминантного гена С, поэтому цветки белые.
Итак, расщепление по фенотипу произошло наполовину” 50 % потомков с пурпурными цветками, 50 % - с белыми.
Задача 2. Растение с белыми цветками, скрещенное с таким же, дает 3/4 потомков с белыми и 1/4 с пурпурными цветками. Каковы генотипы родителей? (Обозначения генов и признаков те же, что и в предыдущей задаче.)
Все гены родительских генотипов в данной задаче не известны, потому что белая окраска цветков обусловлена генами С, Р и их рецессивной гомозиготой - ссрр. Задачу можно решить исходя из анализа пурпурных растений в потомстве.
По условию задачи пурпурная окраска цветков обусловлена наличием в генотипе двух доминантных генов - С и Р. В потомстве есть растения с пурпурными цветками, которые получили по гену С от матери, по гену Р - от отца. В генотипе одного родителя оба этих гена не могут находиться, т. к. цветки их белые. По установленным генам генотипы родителей можно записать так: С_рр и ссР _.
Теперь следует выяснить, в каком состоянии находятся доминантные гены: в гомо- или гетерозиготном. Для этого анализируем характер расщепления в потомстве. Признак окраски цветков расщепился в отношении 3: 1 - от четырех слияний образовались четыре генотипа. Следовательно, родители по генам С и Р гетерозиготны - Ссрр и ссРр.

В результате скрещивания растений с белыми цветами получили 50 % растений с пурпурными цветами и 50 % - с белыми.