Результативность селекции в первую очередь зависит от применяемых методов. К настоящему времени наиболее распространенными являются методы отбора и гибридизации. Кроме того, в последние годы активные работы ведутся по разработке методов полиплоидии, мутагенеза, культуры изолированных клеток и тканей, клеточной и генной инженерии.
Отбор
В основе селекционной работы в любом направлении отбор является наиболее важным и решающим процессом все сорта сельскохозяйственных культур созданы и создаются отбором.
Улучшение древесных и кустарниковых пород путем выявления, отбора и размножения ценных популяций и форм является основным методом лесной селекции.
В лесной селекции выделяют три типа отбора: массовый, групповой и индивидуальный.
Массовый отбор, или отбор, лучших климатических экотипов (отбор по происхождению), является простейшим методом селекции, который положен в основу районирования переброски семян лесных древесных растений. При помощи географических культур установлено большое влияние происхождения семян на рост и качество лесных насаждений сосны, ели, дуба, лиственницы и других видов древесных растений как в пределах естественного ареала, так и при интродукции. На первом этапе искусственного отбора выгодно исходить из местной, или локальной, расы (популяции), которая под действием естественного отбора приобрела полезные приспособления к окружающим условиям.
Групповой отбор лучших местных популяций становится основой лесной селекции. Этот вид отбора высококачественных насаждений, популяций лесных древесных пород называют популяционным отбором.
Индивидуальный отбор – это отбор лучших биотопов, или клонов, а также ценных форм в популяциях и экотипах. Индивидуальный отбор плюсовых деревьев по фенотипу без проверки по потомству соответствует однократному массовому отбору в сельском хозяйстве. Отбор лучших особей в ряду поколений сельскохозяйственных растений получил название непрерывного массового отбора. Разновидностью непрерывного массового отбора является негативный отбор, при котором отбираются не лучшие растения, а удаляют из насаждения худшие особи. В лесоводстве процесс негативного отбора осуществляется рубками ухода и санитарными рубками, при которых вырубаются худшие по селектируемому признаку и больные растения.
Отбор по фенотипу дает хорошие и быстрые результаты в том случае, когда его задачи совпадают с направлением естественного отбора в данных условиях. Например, если в популяциях имеются рано распускающиеся формы и этот признак в данной местности определяет лучшее вызревание побегов и улучшает общий прирост древостоев, то отбор по началу и продолжительности вегетации может быть эффективным. Но в селекции на улучшения качества продукции возможности отбора по фенотипу часто очень ограничены. Таким образом, аналитическая селекция и создание новых форм при помощи других методов значительно облегчаются при совпадении направлений естественного и искусственного отборов. В популяциях лесных древесных растений наблюдаются три основных формы естественного отбора: направленный, стабилизирующий и дизруптивный.
Направленный отбор - из популяции в процессе развития древостоев отмирает один из генотипов с положительным или отрицательным отклонением значения признака от среднепопуляционного. Направленный отбор у лесных древесных пород с отпадом в онтогенезе отстающих по росту особей наиболее характерен для признака продуктивности по высоте.
Стабилизирующий отбор наблюдается при пониженной адаптивности ценности признака, когда в популяции отмирают особи, имеющие генотипы с крайними отклонениями признака (возрастает частота гетерозигот). Например, превалирования прироста гетерозиготных особей в раннем возрасте у ели, березы и других пород.
Дизруптивный отбор – определенные преимущества имеют оба генотипа с крайними отклонениями признака. В зависимости от типа скрещивания между ними возникает различное распределение генотипов.
Основными методами индивидуального отбора являются методы: клоновый отбор, отбор у перекрестноопыляющихся растений и метод педегри.
Индивидуальный отбор в селекции растений, размножаемых вегетативно, называется клоновым. Индивидуальный отбор у перекрестного опыляющихся растений, вследствие постоянного переопыления и расщепления признаков сортов у перекрестноопыляющихся растений нельзя вывести путем однократного индивидуального отбора. Поэтому в селекции таких растений применяется многократный или непрерывный (в каждом поколении) индивидуальный отбор. При этом имеют дело не с отдельным растением, а с семьями.
Индивидуально-семейный отбор заключается в том, что после отбора лучших растений в популяции исходного материала потомство каждого лучшего растения, называемого семьей, размещают изолированно от остальных потомств (Царев и др., 2001).
Гибридизация
Гибридизация – скрещивание между собой двух или большего числа видов, наследственно различающихся по какому-либо признаку или группе признаков.
Гибрид – потомство, полученное в результате скрещивания особей с разной наследственностью.
Скрещивание особей разных форм и сортов, принадлежащих к одному виду, называется внутривидовой гибридизацией.
Скрещивание особей, принадлежащих к разным видам одного рода, разным родам и разным семействам, называется межвидовой, межродовой и межсемейственной гибридизацией.
Гетерозис – увеличение мощности и жизнеспособности гибридов, по сравнению с родительскими формами.
Искусственная гибридизация – контролируемое скрещивание, которое проводится с целью получения семенного потомства, отличающегося хозяйственно-ценными признаками.
При гибридизации:
1) подбирают родительские пары для скрещивания (чтобы каждый из родителей обладал одним из признаков, которые необходимо получить в гибриде)
2) наблюдают за протеканием репродуктивной фазы у родительских особей
3) собирают пыльцу, определяют ее жизнеспособность
4) проводят искусственное опыление
5) наблюдают за развитием гибридных семян
6) собирают гибридные семена, высеивают и выращивают гибридное потомство
7) отбирают среди гибридов экземпляры по селектируемым признакам.
В зависимости от принципа подбора родительских пар различают типы скрещиваний:
Простые – однократные скрещивания между двумя родительскими формами
Парные – однократное скрещивание только между двумя родительскими формами
Диаллельные – каждая испытываемая форма или вид скрещивается со всеми другими формами или видами во всех возможных комбинациях
Реципрокные – каждый из двух форм или видов в одном случае является материнской формой, во втором отцовской
Сложные – если скрещивается более двух родительских форм или гибридное потомство повторно скрещивается с одним из родителей
Множественные (поликросы) – материнское растение опыляется смесью пыльцы нескольких видов
Возвратные (беккроссы) – гибрид повторно скрещивается с одной из родительских форм
Ступенчатые – полученный от простого скрещивания гибрид скрещивается не с родительской формой, а с третьим видом, затем с четвертым
Конвергентные – скрещивание гибридов от материнской и отцовской форм
Межгибридное – объединение наследственности нескольких родителей при скрещивании гибридов от параллельного скрещивания (Любавская, 1985).
Мутагенез
Мутации - это прерывистые, скачкообразные изменения наследственных структур, возникающие под влиянием факторов среды. Изменчивость, обусловленная возникновением мутаций, называется мутационной. Растения с мутировавшими клетками называются мутантами.
Мутации классифицируют по различным признакам. В зависимости от того, на каком уровне они возникают и какие генетические структуры затрагивают, различают мутации генные, хромосомные, геномные, пластидные и плазменные.
Генные мутации представляют собой наследственные изменения, связанные с появлением новых аллелей. Они возникают на уровне нуклеотидов в результате потери или удвоения, изменения порядка чередования или вставки одного или нескольких нуклеотидов в молекулах ДНК в ядре. Так как фиксированных границ между триплетами не существует, все перечисленные изменения в молекулах ДНК приводят к изменению порядка "считывания" наследственной информации, заключенной в этих молекулах.
Хромосомные мутации возникают вследствие разрывов хромосом. Оторванная часть хромосомы (фрагмент) может присоединиться к той же или любой другой хромосоме или элиминируется в процессе деления клетки. Потеря части хромосомы называется делекцией, удвоение одного и того же участка хромосомы - дупликацией, поворот в пределах одной хромосомы какого-то участка ее на 180° получил название инверсия, а присоединение фрагмента к другому месту той же или другой негомологичной хромосомы - транслокации. Перестройки, захватывающие всю хромосому, называются хромосомными, а хроматиду - хроматидными (Котов, 1997).
Геномные мутации связаны с изменением числа хромосом в клетке.
Плазменные мутации представлены изменениями наследственных структур в цитоплазме, а пластидные - в пластидах. Они изучены пока недостаточно полно.
Элементарной единицей мутации считается мутон, равный одному нуклеотиду.
Мутации возникают в природе и могут быть вызваны искусственно. Первые называются спонтанными (естественными), а вторые - индуцированными (искусственными). По своей природе мутации естественного происхождения и полученные искусственно не отличаются.
Мутации, возникающие в гаметах, называются гаметическими, а в клетках тела - соматическими. Если мутировавшая гамета участвует в оплодотворении и образовании зиготы, то все клетки организма, вырастающего из такой зиготы, будут мутировавшими. Соматическая мутация передастся только тем клеткам организма, для которых в качестве исходной материнской служила соматическая мутантная клетка. Таким образом возникает химерный организм, состоящий из клеток с неодинаковой генетической информацией.
По характеру фенотипического проявления различают мутации морфологические, если изменение затрагивает морфологические признаки, физиологические и биохимические. В последних двух случаях изменения касаются физиологических и биохимических процессов. Строго говоря, любые изменения морфологических признаков или функциональных особенностей происходят через изменения биохимических процессов (Котов, 1997).
В большинстве случаев мутации снижают жизнеспособность мутировавшей клетки и организма. Такие мутации называются вредными. Крайним выражением действия вредных мутаций является гибель мутанта. В таких случаях мутация называется летальной. Мутации, не вызывающие заметных сдвигов в жизнеспособности организмов, - нейтральные. Часть мутаций способствует усилению жизнеспособности организмов. Это - биологически полезные мутации. Их нельзя отождествлять с хозяйственно полезными мутациями, вызывающими усиление признаков, используемых человеком для удовлетворения своих нужд. Нередко улучшение "хозяйственного" признака сопровождается ослаблением конкурентной способности организма. Естественный отбор такие мутанты элиминирует, а человек отбирает и разводит в культуре. Любая мутация может наследоваться, так как она возникает в наследственных структурах клеток. Характер наследования зависит от природы мутаций, хотя в основе лежат общие закономерности, связанные с их наследованием.
Факторы, вызывающие изменения в молекулах ДНК или хромосомах, называются мутагенными. Мутагенными свойствами обладают такие физические факторы как ионизирующая радиация, ультрафиолетовые лучи, температурные шоки, лазерное излучение, а также многие химические соединения.
В настоящее время в мутационной селекции наибольшее применение находят ионизирующие излучения, которые по своей природе подразделяются на волновые и корпускулярные. К волновым излучениям относятся ультрафиолетовые лучи (длина волны 2000-4000 а), рентгеновы лучи (0,05 - 10 а) и гамма-лучи (менее 0,05А). К корпускулярным излучениям относятся а-частицы, протоны, нейтроны, дейтроны и др.
В основе оценки чувствительности к мутагену лежит общая закономерность, известная под названием закона Арндта-Шульце. Суть его заключается в том, что слабое воздействие мутагеном способствует стимуляции процессов жизнедеятельности растений. По мере усиления воздействия мутагеном стимулирующий эффект возрастает и достигает своего предела, затем постепенно падает и снижается до нуля, после чего переходит в свою противоположность, то есть начинает подавлять процессы жизнедеятельности и тем больше, чем сильнее действие мутагенов. Подавляющее воздействие мутагенов постепенно возрастает вплоть до полной гибели клеток, органа или всего растения. Схематически закон Арндта-Шульце представлен на примере отзывчивости желудей дуба черешчатого на рентгеново облучение.
Различают стимулирующие, критические, летальные и оптимальные дозы мутагенов. Дозы мутагена определяются мощностью источника излучения или концентрацией его (для химических мутагенов) и продолжительностью воздействия на растения, или экспозицией.
Единицей дозы облучения волновыми излучениями служит рентген, равный количеству излучения, при котором в 1 см 3 сухого воздуха при 00С и давлении 760 мм рт. ст. образуется 2,1*109 дар ионов. Доза нейтронов определяется их количеством, приходящимся на 1 см 2 облучаемой поверхности. 1000 рентген (р) равны 1 килорентгену (кр).
Сравнимость доз облучения достигается при их переводе в дозы поглощения. Единицей дозы поглощения является 1 рад., равный количеству излучений, эквивалентному поглощению 1 граммом вещества энергии в 100 эрг. 1 рад соответствует 1,07 рентгена.
Стимулирующими называются дозы, при которых наблюдается совершенствование процессов жизнедеятельности у обрабатываемых растений в сравнении с контролем, например, повышение всхожести семян, ускорение роста, увеличение урожая и т.д.
Дозы, при которых всхожесть семян составляет около 50% от контроля, а выживаемость - 20-30% от числа всходов, называются критическими.
Дозы, вызывающие гибель обрабатываемого материала, называются летальными, а дозы, при которых на единицу выживаемости растения получается наибольшее количество мутаций, - оптимальными.
При оценке чувствительности растения к мутагенам учитываются такие признаки, как энергия прорастания семян, их всхожесть, выживаемость растений, степень подавления роста, плодовитость и стерильность растения, число и типы хромосомных перестроек в первом митозе в клетках проростков.
Существенные изменения в реакции организма на облучение вносят технические условия обработки: вид излучения, мощность и величина дозы, содержание кислорода при облучении и после него.
Сходная картина наблюдается при обработке растений химическими мугагенами.
Постановка задач при селекции методом индуцированного мутагенеза исходит из направлений селекции и района работы. Растения подбирают, как правило, с минимальным числом отрицательно выраженных признаков, которые подлежат улучшению посредством мутагенеза. В качестве растительного материала для получения гаметических мутаций берут пыльцу, срезанные до споро- или гаметогенеза ветви с пестичными или тычиночными цветками, а также целые растения. Для получения соматических мутаций используют семена, черенки с вегетативными почками, целые растения. Растительный материал подбирают в таком физиологическом состоянии, чтобы при имеющихся в распоряжении селекционера технических возможностях было бы возможно выращивание растений из обработанного мутагенами материала.
Выбор мутагенов зависит от применяемых доз и практической их доступности. В районных центрах и поселках, где есть лечебные учреждения с терапевтической рентгеновской установкой, доступны ультрафиолетовые лучи для работы с пыльцой и каллусными тканями, а также рентгеновы лучи для работы с пыльцой, семенами, черенками. В областных центрах и крупных городах, где есть онкологические отделения, вполне реально использование гамма-лучей кобальтовых пушек. Не является проблемой приобретение в лесничества или лесхозы гупосов (гамма-установок предпосевной обработки семян), химических мутагенов и элементарного оборудования для работы с ними.
Обработка растений корпускулами пока возможна только в ядерных колонках нескольких научно-исследовательских центров. Функционируют международные центры по мутагенезу с гамма-полями. Для выбора рабочих доз обработки растительных объектов необходимо располагать информацией о чувствительности растений к мутагену. Если такой информации нет, то предварительно экспериментальным путем следует установить области стимулирующих, нейтральных, оптимальных, критических и летальных доз. В дальнейшем в целях получения большего количества точковых мутаций следует работать в области стимулирующих и нейтральных доз. Если нужно получить хромосомные и хроматидные мутации, рекомендуется работать в области оптимальных и критических доз.
В работе с химическими мутагенами пользуются 0,00001-0,01%-ми концентрациями растворов. Обязательны меры предосторожности в работе со всеми мутагенами.
Обработанный мутагенами материал используется далее или для скрещивания, или для посадки, прививки, посева. Организуются наблюдения за опытными и контрольными растениям. Ведется журнал селекции, где отмечаются все характеристики исходного материала (вид; внутривидовой таксой; адрес и условия произрастания; какой орган обработан; когда, где и как заготовлен и хранился исходный материал; когда, чем, как обработан; что сделано после обработки; результаты наблюдений).
Перспективные гаметические и соматические мутанты могут быть сразу отобраны. В таких случаях они поступают на сортоиспытание.
Если же наряду с полезными мутанты несут отрицательные изменения, то их скрещивают с ценными видами или формами или повторно обрабатывают мутагенами, а затем уже производят отбор и испытание представляющих интерес мутантов.
Как уже отмечалось, соматические мутации могут возникать не во всех клетках обрабатываемого растения, а только в некоторых из них. В этих случаях растения оказываются химерными. Как правило, измененные клетки делятся медленнее нормальных, что приводит к постепенному вытеснению мутантных тканей нормальными. Это явление получило название соматического отбора» Для выявления скрытых соматических мутаций применяются специальные приемы расхимеривания. У древесных видов расхимеривание можно провести путем одной или многократной посадки опытного растения на пень и последующего отбора мутантных порослевин, черенкования отдельных частей растения, прививки черенков и почек, микроклонального размножения (Котов, 1997).
Для получения мутанта лиственницы сибирской следует взять семена в количестве 50000 шт. и обработать рентгеновым излучением 4000 рентген. В результате облучение половина семян погибла, остальные 25000 семян высеивают и ухаживают за ними до самой смерти. Лучшие мутанты в количестве 5000 шт. отбирают и размножают.
Полиплоидия
Полиплоидией называется биологическое явление, характеризующееся нетипичным числом хромосом в клетках. Растения с нетипичным числом хромосом в клетках называются полиплоидами. Они подразделяются на эуплоиды и анэуплоиды. Эуплоиды - это растения с числом хромосом, кратным основному (гаплоидному) набору. Если, например, у березы основное число хромосом равно 14, то типичными являются гаметы с 14, а соматические клетки с 28 хромосомами. Гаметы с 28, 42, 56 хромосомами будут нетипичными, полиплоидными или нередуцированными. Соматические клетки с 14 хромосомами называются гаплоидными, с 42 - трипловидными, 56 - тетраплоидными и т.д. Если в клетках увеличивается число геномов одного вида, растения называются автополиплоидами. Например, у осины есть автотриплоиды с тремя наборами геномов в клетках (Зn). Число хромосом в них 57, так как в геноме 19 хромосом. Если растения представляют собой гибриды между двумя видами и в клетках содержится не по одному геному каждого вида, а по крайней мере по два генома хотя бы одного вида, они называются аллополиплоидами. Например, среди растений тополя сереющего встречаются триплоиды. В клетках таких растений содержится по три генома, один из которых представлен геномом осины, а два - геномом тополя белого или наоборот.
Растения, в клетках которых содержится число хромосом некратное основному (гаплоидному), называются анэуплоидами. Например, вяз мелколистный имеет 14 хромосом в основном наборе. В степях Северного Казахстана обнаружены растения с 29 хромосомами в соматических клетках. Они имели два набора хромосом (2п=28) и одну добавочную хромосому.
Полиплоиды возникают разными путями. Один из них - нарушение сократительной функции ахроматинового веретена при митозе или мейозе. Вследствие этого хромосомы или хроматиды теряют ориентацию при расхождении к полюсам и в дочерние клетки может отойти число хромосом от 0 до 2п независимо от способа деления (митоз или мейоз), от стадии формирования спор и гамет. Клетки с несбалансированным числом хромосом (не кратным основному) характеризуются слабой жизнеспособностью.
В природе встречаются такие деления, как кариокинез без цитокинеза и удвоение хромосом без последующего деления ядра (эндомитоз). В обоих случаях возникают полиплоидные клетки.
Полиплоиды могут быть и комбинативного происхождения, когда зигота образуется от слияния гамет с разным набором геномов. Слияние гаплоидной и диплоидной гамет дает триплоидную зиготу, двух диплоидных - тетраплоидную и т.п.
Анэуплоиды возникают, как правило, вследствие воздействия на растения мутагенов. Например, в процессе репарации разорванных хромосом возникают ацентрические и дицентрические хромосомы. Первые образуются при объединении фрагментов, не имеющих центромеры. При делении клетки такая хромосома теряется. Дицентрические хромосомы возникают при срастании двух фрагментов с центромерами, которые образуют в анафазе так называемые мосты и далее или снова разрываются, или превращаются в кольцевые хромосомы, которые в клетке не элиминируются (Котов, 1997).
Для получения полиплоида лиственницы сибирской рекомендуется взять 20000 семян, произвести намачивание семян в 0,1%-ном растворе колхицина. Продолжительность действия 18 часов. После этого промывают проточной водой. Затем высеивают, после появления всходов обследуют на появление полиплоидов. Полиплоиды оставляют на контроль, а остальные оставляют и сравнивают с контролем.
Генная инженерия
За последние 10-15 лет были созданы принципиально новые методы манипулирования с нуклеиновыми кислотами in vitro, на основе которых зародился и бурно развивается новый раздел молекулярной биологии и генетики - генная инженерия. Принципиальное отличие генной инженерии от использовавшихся ранее традиционных приемов изменения генотипа (например, создания полиплоидных форм растений) состоит в том, что она дает возможность конструировать функционально активные генетические структуры in vitro в форме рекомбинантных ДНК. Понятия «генная» и «генетическая» инженерия часто употребляют как синонимы, хотя последнее является более широким и включает манипулирование не только отдельными генами, но и с более крупными частями генома. Работа по переделке генотипа животных или растений с помощью скрещиваний ограничены пределами вида либо близких в видовом отношении форм. Напротив, генная инженерия, как будет показано ниже, стирает межвидовые барьеры, обеспечивая возможность создания организмов с новыми, в том числе и не встречающимися в природе, комбинациями наследственных свойств. Генная, инженерия представляет собой совокупность методов, позволяющих не только получать рекомбинантные ДНК из фрагментов геномов разных организмов, но и вводить такие рекомбинантные молекулы в клетку, создавая условия для экспрессии в ней введенных, часто совершенно чужеродных генов. Таким образом, в этом случае исследователь оперирует непосредственно с генами, причем их перенос может не зависеть от таксономического родства используемых организмов. Эта особенность генной инженерии представляет ее главное отличие от ранее использовавшихся приемов изменения генотипа.
Первенствующую роль в формировании генной инженерии сыграла генетика микроорганизмов, идеи и методы, разработанные молекулярной генетикой и химией нуклеиновых кислот.
Выполнение любой генно-инженерной программы включает необходимость получения фрагментов ДНК, несущих нужный ген, объединение их in vitro с век-торными молекулами, способными обеспечить доставку гена в организм реципиента, создание условий для стабильного наследования и эффективной экспрессии перенесенного гена. Осуществление такой работы определяется крупными достижениями в области генетики и химии нуклеиновых кислот. К важнейшим из них относятся:
1) открытие явления рестрикции-модификации ДНК, в результате которого были выделены необходимые ферменты - рестриктазы для получения специфичных фрагментов ДНК;
2) создание методов химического и химико-ферментативного синтеза генов;
3) выявление векторных молекул ДНК, способных перенести в клетку чужеродную ДНК и обеспечить там экспрессию, соответствующих генов;
4) разработка методов объединения фрагментов ДНК из разных источников;
5) разработка методов трансформации у различных организмов и отбора клонов, несущих рекомбинантные ДНК.
Совокупность этих достижений и составляет сущность методологии генной инженерии.
Не менее важное значение имеет генная инженерия в качестве мощного инструмента фундаментальных исследований. С ее помощью изучают строение различных геномов, отдельных генов и кодируемых ими продуктов. Генная инженерия помогла раскрыть экзонинтронную организацию эукариотических генов, позволяла понять суть явления непостоянства генома, связанного с присутствием мигрирующих генетических элементов у про- и эукариот, открыла принципиально новые возможности для изучения молекулярных основ онтогенеза, наследственных заболеваний, эволюционного происхождения различных организмов. В значительной мере этим успехам генной инженерии способствовало создание банков (или библиотек) генов отдельных организмов, резко облегчающих стратегию поиска индивидуальных генов, исследование их структуры и функции. Получение танков генов включает выделение тотальной ДНК, фрагментацию ее с помощью рестриктаз, присоединение полученных фрагментов к векторным молекулам (плазмидного или фагового происхождения) и введение рекомбинантных ДНК в реципиентные бактерии. Эта техника позволяет получить набор клонов бактерий или щтоков гибридных фагов, различающихся по включенным фрагментам ДНК. Необходимые исследователю гены отбирают из таких банков с помощью специально разработанных генетических, биохимических, радиоизотопных и иммунологических методов. Потенциальные возможности генной инженерии действительно очень велики, и их реализация в полной мере дело сегодняшнего дня и ближайшего будущего (Котов, 1997).
С помощью генной инженерии лиственнице сибирской можно пересадить ген лимонника китайского, тем самым создать стелющуюся форму лиственницы. Затем провести наблюдение в течение всей жизни.
Основные методы селекции растений
Классическими методами селекции растений были и остаются гибридизация и отбор.
Размещено на реф.рф
Различают две основные формы искусственного отбора: массовый
и индивидуальный
.
1. Массовый отбор применяют при селекции перекрестноопыляемых растений, таких, как рожь, кукуруза, подсолнечник. При этом выделяют группу растений, обладающих ценными признаками. В этом случае сорт представляет собой популяцию, состоящую из гетерозиготных особей, и каждое семя даже от одного материнского растения обладает уникальным генотипом. С помощью массового отбора сохраняются и улучшаются сортовые качества, но результаты отбора неустойчивы в силу случайного перекрестного опыления.
2. Индивидуальный отбор эффективен для самоопыляемых растений (пшеницы, ячменя, гороха). В этом случае потомство сохраняет признаки родительской формы, является гомозиготным и принято называть чистой линией . Чистая линия - потомство одной гомозиготной самоопыленной особи. У любой особи тысячи генов, и так как происходят мутационные процессы, то абсолютно гомозиготных особей в природе практически не бывает. Мутации чаще всего рецессивны. Под контроль естественного и искусственного отбора они попадают только тогда, когда переходят в гомозиготное состояние.
3. Естественный отбор в селекции играет определяющую роль. На любое растение в течение всей его жизни действует целый комплекс факторов окружающей среды, и оно должно быть устойчивым к вредителям и болезням, приспособлено к определенному температурному и водному режиму.
4. Инбридинг используют при самоопылении перекрестноопыляемых растений , к примеру, для получения чистых линий кукурузы. При этом подбирают такие растения, гибриды которых дают максимальный эффект гетерозиса - жизненной силы, образуют початки более крупные, чем початки родительских форм. От них получают чистые линии - на протяжении ряда лет, производят принудительное самоопыление - срывают метелки с выбранных растений и, когда появляются рыльца пестиков, их опыляют пыльцой этого же растения. Изоляторами предохраняют соцветия от попадания чужой пыльцы. У гибридов многие рецессивные неблагоприятные гены при этом переходят в гомозиготное состояние, и это приводит к снижению их жизнеспособности, к депрессии. Далее скрещивают чистые линии между собой для получения гибридных семян, дающих эффект гетерозиса.
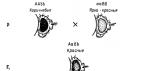
Эффект гетерозиса объясняется двумя основными гипотезами. Гипотеза доминирования предполагает, что эффект гетерозиса зависит от количества доминантных генов в гомозиготном или гетерозиготном состоянии. Чем больше в генотипе генов в доминантном состоянии - тем больший эффект гетерозиса, и первое гибридное поколение дает прибавку урожая до 30% (рис. 339).
ААbbCCdd x aaBBccDDГипотеза сверхдоминирования объясняет явление гетерозиса эффектом сверхдоминирования: иногда гетерозиготное состояние по одному или нескольким генам дает гибриду превосходство над родительскими формами по массе и продуктивности. Но начиная со второго поколения эффект гетерозиса затухает, так как часть генов переходит в гомозиготное состояние.
АА 2Аа аа
5. Перекрестное опыление самоопылителей дает возможность сочетать свойства различных сортов. Рассмотрим, как это практически выполняется при создании новых сортов пшеницы. У цветков растения одного сорта удаляются пыльники, рядом в банке с водой ставится растение другого сорта͵ и растения двух сортов накрываются общим изолятором. В результате получают гибридные семена, сочетающие нужные селекционеру признаки разных сортов.
6. Очень перспективен метод получения полиплоидов, у растений полиплоиды обладают большей массой вегетативных органов, имеют более крупные плоды и семена. Многие культуры представляют из себяестественные полиплоиды: пшеница, картофель, выведены сорта полиплоидной гречихи, сахарной свеклы.
7. Отдаленная гибридизация - скрещивание растений, относящихся к разным видам. Но отдаленные гибриды обычно стерильны, так как у них нарушается мейоз (два гаплоидных набора хромосом разных видов не конъюгируют), и не образуются гаметы.
В 1924 году советский ученый Г.Д.Карпеченко получил плодовитый межродовой гибрид. Он скрестил редьку (2n = 18 редечных хромосом) и капусту (2n = 18 капустных хромосом). У гибрида в диплоидном наборе было 18 хромосом: 9 редечных и 9 капустных, но при мейозе редечные и капустные хромосомы не конъюгировали, гибрид был стерильным.
С помощью колхицина Г.Д.Карпеченко удалось удвоить хромосомный набор гибрида, полиплоид стал иметь 36 хромосом, при мейозе редечные (9 + 9) хромосомы конъюгировали с редечными, капустные (9 + 9) с капустными. Плодовитость была восстановлена. Таким способом были получены пшенично-ржаные гибриды (тритикале), (рис. 341) пшенично-пырейные гибриды и др.Размещено на реф.рф
Виды, у которых произошло объединение разных геномов в одном организме, а
затем их кратное увеличение, называются аллополиплоидами.
8. Использование соматических мутаций применимо для селекции вегетативно размножающихся растений, что использовал в своей работе еще И.В.Мичурин. С помощью вегетативного размножения можно сохранить полезную соматическую мутацию. Вместе с тем, только с помощью вегетативного размножения сохраняются свойства многих сортов плодово-ягодных культур.
9. Экспериментальный мутагенез основан на открытии воздействия различных излучений для получения мутаций и на использование химических мутагенов. Мутагены позволяют получить большой спектр разнообразных мутаций, сейчас в мире созданы более тысячи сортов, ведущих родословную от отдельных мутантных растений, полученных после воздействия мутагенами.
Многие методы селекции растений были предложены И.В.Мичуриным. С помощью метода ментора И.В.Мичурин добивался изменения свойств гибрида в нужную сторону. К примеру, в случае если у гибрида нужно было улучшить вкусовые качества, в его крону прививались черенки с родительского организма, имеющего хорошие вкусовые качества; или гибридное растение прививали на подвой, в сторону которого нужно было изменить качества гибрида. И.В.Мичурин указывал на возможность управления доминированием определенных признаков при развитии гибрида. Для этого на ранних стадиях развития крайне важно воздействие определенными внешними факторами. К примеру, в случае если гибриды выращивать в открытом грунте, на бедных почвах, повышается их морозостойкость.
Основные методы селекции растений - понятие и виды. Классификация и особенности категории "Основные методы селекции растений" 2017, 2018.
Селекция — наука о методах создания и улучшения существующих сортов растений, пород животных и штаммов микроорганизмов.
Она использует научные открытия в области молекулярной биологии, биохимии и генетики для изменения наследственности живого организма.
Что входит в теоретическую базу селекции?
- основные законы генетики, наследственности и изменчивости, знания о структуре гена, молекулярные основы наследственности;
- изучение исходного материала, знание генетики основных видов, сортов, пород и штаммов;
- изучение влияния среды на проявление генотипа;
- знание форм искусственного отбора, выявление необходимых признаков и закрепление их у организмов;
- методы клеточной и генной инженерии.
Задачи селекции
- повышение урожайности сортов растений и продуктивности пород животных и штаммов микроорганизмов
- создание устойчивых к заболеваниям и климатическим условиям сортов и пород
- получение сортов, пород и штаммы, пригодных для механизированного или промышленного выращивания и разведения
Сорт, порода, штамм
— популяции организмов, искусственно
созданные человеком, характеризующиеся определенным генофондом, наследственно закрепленными признаками, определенной продуктивностью и .
Основные методы селекции растений


Массовый отбор — не проверяет генотип каждой особи, поэтому результаты не очень устойчивы;
Индивидуальный отбор — специально увеличивают количество гомозиготных особей; идет выделение чистых линий.
В селекции растений возможно получение потомства путем, чтобы сохранить полученные признаки неизменными.
Методы селекции животных

Факторы, коорые необходимо учитывать при селекции животных:
- у животных значительно меньше потомство, чем у растений, при этом каждый организм имеет хозяйственную ценность;
- половая зрелость наступает поздно;
- нет быстрой смены поколений;
- отсутствует вегетативное размножение и самооплодотворение
- Используется только размножение ;
- размножение происходит реже, чем у растений (особи должны еще достичь полового созревания;
- у животных инбридинг приводит к ослаблению гибридов, уменьшению жизнеспособности ;
- при неродственном скрещивании — наоборот, жизнеспособность гибридов увеличивается ;
- при массовом отборе животных руководствуются стабилизирующим отбором ;
Методы селекции микроорганизмов

Микроорганизмы используются в медицине для получения антибиотиков, гормонов, витаминов и т.п., в промышленности — для производства пестицидов, топлива, очистки воды и т.д, в сельском хозяйстве — для получения кормовых белков, аминокислот, сахарозаменителей, органических кислот и т.д.
Несколько определений:
Трансгенные — виды растений, в которых успешно функционируют гены, пересаженные из других видов растений и животных.
Гетерозис — увеличение жизнеспособности гибридов за счет наследования определенного набора аллелей различных генов от своих разнопородных родителей.
Иннбридинг — близкородственное скрещивание, приводящее к появлению гомозиготных организмов.
Полиплойдия — набор хромосом больше, чем 2n — широко распространена в природе. Высокая степень полиплойдности свойтсвенна простейшим. Среди многоклеточных редка и характерна для видов, утративших нормальный половой процесс — гермафродитов и видов, для которых характерен .
Доместикация — процесс изменения диких животных или растений, при котором на протяжении многих поколений они содержатся человеком генетически изолированными от дикой формы и подвергаются искуственному отбору
Искусственный отбор - выбор человеком наиболее ценных в хозяйственном или декоративном отношении особей животных и растений для получения от них потомства с желаемыми свойствами. Предшественник и основной метод современной селекции.
Давайте сравним искусственный и естественный отбор:
| Естественный отбор | Искусственный отбор (селекция) |
|
Производится природой (условиями среды обитания: биотические и абиотические факторы |
Производится человеком |
|
Признаки:
|
Признаки:
|
|
Формы отбора: движущий стабилизирующий |
Формы отбора: массовый индивидуальный методический стихийный |
|
Результат: новые виды |
Результат: новые породы и сорта |
Выдающиеся ученые — селекционеры
Из отечественных ученых особенности селекции изучали — человека, заложившего основы российской генетики. В школе проходят его закон гомологических рядов:
«изменчивость близких по происхождению родов и видов растений осуществляется общим (параллельным) путем. Генетически близкие роды и виды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других родственных видов и родов»
p style=»text-align: justify;»>В своих экспедициях Вавилов собрал богатейшую коллекцию культурных растений, нашел родственные связи между ними, предсказал возможные для выведения ранее неизвестные, но заложенные генетически свойства этих культур. Он обнаружил существование районов с максимальной концентрацией видов, разновидностей и сортов определенных культурных растений, а также то, что эти районы связаны с местами древнейших цивилизаций.
Необходимо знать основные центры происхождения культурных растений

И, конечно, — слово «мичуринец» стало нарицательным. Очень много сортов растений названо в его честь.
- тренировка по теме — вопросы ОГЭ
- вопросы ЕГЭ по теме
Селекцией растений называется наука, изучающая способы создания новых и усовершенствования имеющихся сортов культурных растений с важными в практике признаками. Различают несколько основных методов селекции: отбор, гибридизация, мутагенез и полиплоидия .
Искусственный отбор – основа селекционного процесса. В комплексе с генетическими методиками отбор дает возможность создавать сорта растений с заранее предопределенными особенностями. Отбор может быть массовым и индивидуальным.
Массовый отбор представляет собой выделение ряда экземпляров по внешним признакам без проверки их генотипа. Хорошие результаты в данном случае отмечаются при высоком коэффициенте наследуемости нужных свойств. Данный вид отбора эффективен относительно качественных особенностей, контролируемых одним либо несколькими генами, и редко дает положительные результаты относительно полигенных качеств с низкими коэффициентом наследования. В этом случае востребован индивидуальный (методический) отбор.
При проведении индивидуального отбора (по генотипу) оценивают потомство каждого отдельно взятого растения в ряду нескольких поколений с учетом контроля особенностей, важных для селекционера. Впоследствии оставляют только те экземпляры растений, которые дали максимальное количество потомков с ценными качествами. Благодаря этому, становится возможным оценивать наследственные свойства отдельно взятых особей и способность передавать их следующим поколениям.
Более эффективной считается методика сочетания отбора с некоторыми видами скрещивания (гибридизации) . Все методы гибридизации можно подразделить на аутбридинг (неродственное, или межсортовое скрещивание) и инбридинг (близкородственное, или внутрисортовое скрещивание). Инбридинг аналогичен самоопылению у растений, что приводит к увеличению гомозиготности. Для так называемых чистых линий растений характерно не только наличие ценных признаков, но и сниженная жизнеспособность, что обусловлено переходом в гомозиготное состояние всех вредных рецессивных мутаций. Если чистые линии продолжать скрещивать между собой, то появляется эффект гетерозиса. Благодаря аутбридингу, внутри одного сорта поддерживаются ценные свойства и улучшаются в следующих поколениях гибридов за счет повышения уровня гетерозиготности потомков и гетерогенности популяции.
Селекционерами часто применяется метод полиплоидии и отдаленной гибридизации при создании новых сортов растений, в частности метод автополиплоидии. В ходе этого процесса происходит увеличение размеров клеток из-за увеличения количества хромосом. Также повышается стойкость растений к воздействию вредных микробов (вирусов, бактерий, грибков), неблагоприятных физических и химических факторов. Доказано, что полиплоидные организмы являются более жизнеспособными, чем диплоидные. 80% ныне существующих культурных растений полиплоидные. К ним относятся зерновые культуры, овощные и плодово-ягодные, лекарственные, технические и декоративные растения, дающие большие урожаи, по сравнению с диплоидными аналогами.
В основе аллополиплоидии лежит метод отдаленной гибридизации – скрещивания экземпляров растений, принадлежащих к разным видам, иногда родам. Так, были получены межвидовые гибриды-полиплоиды редьки и капусты, пшеницы и ржи, обладающих высокой урожайностью, неприхотливостью и устойчивостью ко многим вредным факторам и болезням.
 В селекции растений часто применяют спонтанные мутации
. Так, при естественной мутации экземпляра желтого люпина было получено несколько сортов сладкого люпина, пригодного в качестве корма скота, тогда как исходный вид, содержащий алкалоиды, скот не поедал. Огромное количество мутантов характерно для плодовых культур. Они описаны как новые сорта или существуют в гибридизации с другими формами.
В селекции растений часто применяют спонтанные мутации
. Так, при естественной мутации экземпляра желтого люпина было получено несколько сортов сладкого люпина, пригодного в качестве корма скота, тогда как исходный вид, содержащий алкалоиды, скот не поедал. Огромное количество мутантов характерно для плодовых культур. Они описаны как новые сорта или существуют в гибридизации с другими формами.
В настоящее время во многих научных лабораториях ряда стран проводятся работы по получению индуцированных мутаций. Такие мутанты были выделены с помощью воздействия физических факторов (например, рентгеновского излучения) у некоторых злаковых культур. Они отличаются рядом ценных свойств: повышенной урожайностью, увеличением размера семян и др.
Селекция - наука об улучшении отдельных качеств животных и растений, необходимых человеку, а также о выведении новых сортов растений, пород животных, штаммов микроорганизмов. Для создания культурных сортов используют методы селекции растений.
Селекция
Большинство растений, которые современное человечество употребляет в пищу, является продуктом селекции (картофель, томат, кукуруза, пшеница). На протяжении нескольких веков люди культивировали дикие растения, переходя от собирательства к земледелию.
Направлениями селекции являются:
- высокая урожайность;
- питательность растений (например, содержание белка в пшенице);
- улучшенный вкус;
- устойчивость культур к погодным условиям;
- скороспелость плодов;
- интенсивность развития (например, «отзывчивость» на удобрения или полив).

Рис. 1. Сравнение дикой и сельскохозяйственной кукурузы.
Селекция решила проблемы с нехваткой пищи и продолжает развиваться, внедряя методы генной инженерии. Селекционеры не только улучшают вкус и повышают питательность растений, но и делают их полезными, насыщенными витаминами и химическими элементами, важными для метаболизма.
Для успешной селекции необходимо понимать закономерности наследования признаков, особенности влияния среды, морфологическое строение и способы размножения культивируемых растений.
Методы
Основными методами селекции являются:
ТОП-4 статьи которые читают вместе с этой
- искусственный отбор - выбор человеком наиболее ценных культур для селекции;
- гибридизация - процесс получения потомства от скрещивания разных генетических форм;
- искусственный мутагенез - внесение изменений в ДНК.
Искусственный отбор включает в себя два вида - индивидуальный (по генотипу) и массовый (по фенотипу). В первом случае важны конкретные качества растений, во втором - отбирают наиболее приспособленные особи.
Гибридизация бывает двух видов:
- внутривидовая или близкородственная - инбридинг ;
- отдалённая (межвидовая) - аутбридинг .
Классические методы селекции растений описаны в таблице.
|
Метод |
Суть |
Примеры |
|
Индивидуальный отбор |
Проводят по отношению к самоопыляемым растениям. Выведение единичных особей с нужными качествами и получение от них улучшенного потомства |
Пшеница, ячмень, горох |
|
Массовый отбор |
Проводят по отношению к перекрестноопыляемым растениям. Растения скрещиваются массово. Из полученного потомства отбирают лучшие экземпляры и снова проводят скрещивание. Может повторяться до тех пор, пока не будут выведены нужные качества растений |
Подсолнечник |
|
Инбридинг |
Происходит при самоопылении перекрёстноопыляемых растений. В результате получают чистые (гомозиготные) линии, чтобы закрепить полученный признак. Наблюдается снижение жизнеспособности (инбредная депрессия), т.к. потомки постепенно переходят в гомозиготное рецессивное состояние |
Сорта груш, яблонь |
|
Аутбридинг |
Скрещиваются разные виды, потомки обычно стерильны, т.к. при скрещивании нарушается мейоз, не образуются гаметы. В первом поколении наблюдается эффект гетерозиса - превосходство потомков над родительскими формами за счёт образования гетерозиготных генов. Чем отдалённее в родстве родители, тем ярче проявляется гетерозис |
Гибриды пшеницы и ржи (тритикале), смородины и крыжовника (йошта) |
|
Мутагенез |
Подвергают растения ионизирующему, лазерному излучению, химическому или биологическому воздействию, в результате чего возникают мутации. Чаще всего таким способом вырабатывают устойчивость к заболеваниям и вредителям. Метод усовершенствовала генная инженерия - нужный ген можно «включить» или «выключить» вручную без потери других полезных признаков |
Сорта пшеницы |

Рис. 2. Примеры гибридов.
Неудачный опыт селекции - борщевик Сосновского. Растение культивировалось в качестве корма для скота. Однако впоследствии выяснилось, что новый борщевик легко проникает в экосистемы, вытесняя естественные растения, а также содержит вещества, повышающие чувствительность к ультрафиолету. Попав на кожу, сок вызывает ожог на солнце.